Bases fisiopatológicas de la epilepsia del
lóbulo temporal: estudios en humanos y animales
J. Pastor a, Y.G. Uzcátegui c, B. Gal-Iglesias c,d, G.J. Ortega b, R.G. Sola b, L. Menéndez de la Prida c THE PATHOPHYSIOLOGICAL FOUNDATIONS OF TEMPORAL-LOBE EPILEPSY: STUDIES IN HUMANS AND ANIMALSSummary. Introduction. Temporal lobe epilepsy (TLE) is the most frequent form of pharmaco-resistant epilepsy in human. Research using material from TLE patients undergoing surgery and animal models has significantly increased in the last decade. Development. We review recent findings obtained over the last years from electrophysiological and anatomical studies in human and animal models of TLE. Data suggest a large heterogeneity and inter-individual variability depending on the model and the system under study. However, a common principle that appears to underlie the epileptic condition is the reorganization of excitation and inhibition resulting in hyperexcitability. Recent research combining in vitro electrophysiology together with depth recordings in vivo and new analytical methodologies is also discussed. Conclusions. A multidisciplinary approach using both human and animal models can help to fill gaps in our knowledge and to provide unique insights into the pathophysiology of TLE. [REV NEUROL 2006; 42: 663-73] Key words. EEG. Intracellular recordings. Kindling. Membrane receptors. Mesial sclerosis. Microelectrodes. Temporal lobe epilepsy. INTRODUCCIÓN
crisis [8]. El uso de microelectrodos extracelulares ayudó a es-
Los estudios electrofisiológicos sobre la epilepsia en humanos
tablecer la relación entre potenciales sinápticos –de cinética mu-
se iniciaron en el primer tercio del siglo pasado, con el desarro-
cho más lenta que los potenciales de acción– y las ondas obser-
llo de la electroencefalografía (EEG). En 1929, el psiquiatra Hans
vadas en el EEG [9-11]. La utilización de registros intracelu-
Berger (1873-1941), profesor en el Hospital Universitario de
lares, fundamentalmente en modelos animales de hipocampo,
Jena (Alemania), hizo público el hallazgo de que era posible
permitió demostrar la relación entre las posdescargas en el EEG
medir sobre la piel del cráneo en humanos la actividad eléctrica
y los cambios en el potencial transmembrana [12-14], especial-
cerebral [1]. A partir de entonces, el uso del EEG en el diagnós-
mente debido a la suma de potenciales postsinápticos excitato-
tico y tratamiento de la epilepsia, así como en el estudio de su
rios e inhibitorios [15]. Con posterioridad se pudo comprobar
fisiopatología, se extendió con rapidez. Con el uso de tubos de
que la actividad intracelular, tanto durante la actividad interictal
vacío en lugar de galvanómetros, Gibbs et al demostraron la pre-
como durante las crisis, presentaba un elemento común consis-
sencia de complejos punta-onda interictales durante las crisis de
tente en una despolarización extrema y limitada en el tiempo
ausencia [2], mientras que trabajos subsiguientes demostraron la
conocida como ‘descarga paroxística despolarizante’ (paroxys-
presencia de puntas epileptógenas en epilepsias focales [3,4]. En
mal depolarization shift, PDS), característica esencial del foco
la actualidad, el desarrollo de nuevas técnicas analíticas y de
epileptógeno [16-19]. Estas descargas –y las correspondientes
registro del EEG ha motivado un espectacular desarrollo del de-
puntas del EEG– pueden generarse a partir de columnas cortica-
nominado ‘EEG cuantitativo’, un aspecto altamente técnico de
les en torno a los 2 mm, tanto en el córtex de gato [20] como en
la neurofisiología, que presumiblemente puede arrojar luz sobre
numerosos aspectos diagnósticos y fisiopatológicos de la epi-
Desde estos trabajos iniciales hasta el momento actual, los
lepsia y de otras patologías neurológicas y psiquiátricas [5-7].
estudios electrofisiológicos, tanto en modelos animales como
La investigación en neurofisiología se ha dirigido, en gran
utilizando tejido humano in vitro, han experimentado un impor-
medida, hacia la comprensión de la actividad celular responsa-
tante desarrollo. En este trabajo se pretende revisar algunos de
ble de las manifestaciones eléctricas interictales y durante las
los datos y teorías más aceptadas acerca de los mecanismos fi-siopatológicos de la epilepsia del lóbulo temporal (ELT) y sus
Aceptado tras revisión externa: 24.03.06.
manifestaciones electrofisiológicas, tanto en modelos animales
Sección de Neurofisiología Clínica. b Servicio de Neurocirugía. HospitalUniversitario La Princesa. c Departamento de Neurobiología-Investigación. Hospital Ramón y Cajal. d Departamento de Ciencias Morfológicas y Fisio-logía. Universidad Europea de Madrid. Madrid, España.MODELOS ANIMALES DE EPILEPSIA Correspondencia: Dr. Jesús Pastor Gómez. Servicio de Neurofisiología Clí-DEL LÓBULO TEMPORAL nica. Hospital Universitario La Princesa. Diego de León, 62. E-28006 Ma-
Los dos modelos más usados para estudiar la ELT en animales
drid. Fax: +34 914 013 582. E-mail: [email protected]
de laboratorio son el kindling y el modelo de estado epiléptico. Agradecimientos. Al Dr. Norberto García-Cairasco, por sus comentarios so-bre este manuscrito
Aunque ambos inducen un estado epiléptico crónico, el procesode epileptogénesis difiere sustancialmente, así como diferentes
Este trabajo ha contado con financiación de los siguientes proyectos: MECBFI2003-04305 (JP y LMP), Comunidad de Madrid GR/SAL/0131/2004(LMP) y Ministerio de Sanidad FIS 04/216 (GJO).
El fenómeno de kindling, descubierto por Goddard en los
años sesenta [22], consiste en la estimulación repetida y perió-
dica de estructuras del sistema límbico (típicamente de la amíg-
haya visto que el foco se desplaza hacia la corteza piriforme en
dala, la corteza o el hipocampo), que da lugar a la aparición de
las ratas epilépticas [36]. La corteza perirrinal participa de di-
crisis epilépticas secundariamente generalizadas. Durante una
ferentes bucles de actividad recurrente con el hipocampo y la
fase inicial, la estimulación eléctrica induce descargas focales
corteza entorrinal, lo que facilita la iniciación de actividad
con escasa manifestación clínica. Sin embargo, en sesiones con-
reverberante en los circuitos de asociación y la generación de
secutivas, comienzan a registrarse posdescargas en el EEG de la
descargas paroxísticas [37]. Trabajos del grupo de McIntyre
corteza, el hipocampo y la amígdala, alteraciones en la morfolo-
han mostrado que la parte más anterior de la corteza perirrinal,
gía de las puntas y manifestaciones comportamentales más
en el borde con la corteza insular, es crítica para el desarrollo
complejas, como automatismos orofaciales y de las patas delan-
teras, y levantamiento con pérdida de equilibrio, hasta progresar
Al igual que lo descrito en tejido humano, las posdescargas
a crisis tonicoclónicas secundariamente generalizadas [22,23].
corticales se bloquean con antagonistas de los receptores NMDA,
En este modelo, las crisis espontáneas no se desarrollan a no ser
como el AP-5, lo que sugiere la importancia de la transmisión
que se haya realizado un gran número de estimulaciones (mode-
glutamatérgica en su generación [39]. Recientemente se ha des-
lo over-kindling) [24,25]. Se reconoce el kindling como un mo-
crito la supresión del efecto kindling en ratones transgénicos
delo progresivo de ELT en el cual no existe necesariamente da-
que sobreexpresan la subunidad NR2D del receptor NMDA
ño morfológico. Este modelo se usa en la evaluación de fárma-
[40], la cual dota de una menor afinidad por glutamato. Estu-
cos antiepilépticos, pues se ha demostrado que los compuestos
dios moleculares de la composición de estos receptores en epi-
que son efectivos en el tratamiento de la ELT en el hombre, in-
lepsia inducida por kindling han mostrado incrementos de la
hiben las crisis inducidas por kindling [26].
subunidad NR1 en las cortezas temporal y frontal, decrementos
En el modelo de estado epiléptico se utilizan diferentes agen-
en la corteza piriforme [41] y ausencia de alteraciones en el
tes convulsionantes (inyección sistémica o intracerebral), como
hipocampo [42]. Sin embargo, en el modelo de estado epilépti-
el ácido kaínico y la pilocarpina, para inducir una fase aguda de
co, se ha descrito el incremento de las subunidades NR1 y NR2
estado epiléptico caracterizado por crisis tonicoclónicas de ori-
en el giro dentado del hipocampo [43]. Aunque los resultados
gen límbico que no ceden. Una vez superada esta fase, que debe
son ambiguos en diferentes modelos de kindling y estado epi-
interrumpirse farmacológicamente, los animales desarrollan un
léptico, la importancia de los receptores NMDA en la epilepto-
período libre de crisis –fase latente–, seguido de la emergencia
génesis está más que contrastada. Es interesante que inyeccio-
brusca de crisis recurrentes espontáneas –fase crónica– semanas
nes focales repetidas de NMDA en la amígdala produzca un
más tarde [27,28]. En contraste con el kindling, las alteraciones
efecto kindling que se bloquea con la aplicación simultánea de
anatómicas presentes en el hipocampo de las ratas sometidas al
antagonistas como el AP7 [44]. De hecho, antagonistas no com-
modelo de estado epiléptico resultan similares a las descritas en
petitivos de NMDA como el MK801 han mostrado claros efec-
el humano, por lo que éste constituye el modelo más aceptado
tos antiepileptógenos tanto en los modelos de kindling como de
de ELT. No obstante, es importante reconocer las diferencias de
estado epiléptico inducido por pilocarpina [45,46].
cada modelo con el caso humano y utilizarlos de manera com-
Los receptores AMPA y kainato participan de la modula-
plementaria para comprender los diferentes aspectos fisiopato-
ción de las posdescargas, aunque su papel en la epileptogénesis
es aún objeto de estudio. Inyecciones locales de los antagonistasCNQX y GYKI52466 suprimen las crisis sólo transitoriamente
Alteraciones en regiones corticales
en el modelo de kindling del hipocampo [47]. Sin embargo, el
Estudios electrofisiológicos y metabólicos, utilizando mapeo de
NBQX se muestra eficaz en suprimir las descargas cuando se
2-deoxiglucosa, demuestran la participación de la formación
estimula la amígdala [48], lo que sugiere la participación de
del hipocampo, la amígdala y las cortezas perirrinal y piriforme
diferentes circuitos de iniciación y relevo según el modelo utili-
en las crisis inducidas por ambos modelos de ELT [29,30]. Uti-
zado. Otro aspecto importante tiene relación con la dinámica de
lizando inmunohistoquímica contra Fos –una de las proteínas
expresión de los diferentes receptores. En la corteza piriforme
codificadas por genes de expresión temprana–, se ha visto que
se ha demostrado un decremento transitorio de GluR2, subuni-
los patrones de activación durante las fases iniciales se restrin-
dad del receptor AMPA que lo hace impermeable a Ca2+ sólo a
gen a estas estructuras, además de los núcleos de la línea media
las 24 horas del inicio del kindling [49].
del tálamo y el núcleo accumbens [31]. Electrofisiológicamen-
Curiosamente, el test de pares de pulsos en la corteza piri-
te, las cortezas perrininal y piriforme están entre las primeras
forme de ratas sometidas a kindling sugiere un incremento de la
estructuras que generan posdescargas. Además, el umbral de ex-
inhibición [50]. Esto podría contradecir la capacidad de estos
citabilidad disminuye rápidamente con episodios sucesivos de
circuitos para producir posdescargas. Sin embargo, se ha descri-
crisis focales [32]. De hecho, el kindling en la corteza piriforme
to que una inhibición exacerbada proporciona un mecanismo
sigue un patrón rápido de establecimiento que se ha utilizado
para coordinar la actividad aumentando la sincronización en
circuitos glutamatérgicos aberrantes [51]. Por otro lado, estu-
El estudio in vitro de rodajas de corteza perirrinal ha mos-
dios inmunohistológicos han mostrado que el kindling de amíg-
trado que las descargas de tipo interictal se mantienen en la
dala provoca una reducción del número de neuronas gabérgicas
corteza aislada, lo que sugiere que el sitio de iniciación es local
en áreas discretas de la corteza piriforme [52]. Esto sugiere que
[34]. Este tipo de actividad eléctrica puede registrarse en mate-
las alteraciones pueden ser locales y específicas. Es posible que
rial de animales controles, lo que denota que estos circuitos son
el aumento de la inhibición en las cortezas perirrinal y piriforme
especialmente epileptógenos. Celularmente, las neuronas de las
se deba a una respuesta compensatoria a la excesiva activación
capas II y III de la corteza piriforme de ratas epilépticas mues-
de estas estructuras durante las fases iniciales del estableci-
tran tasas de descarga mayores comparadas con controles [35].
miento del modelo. Otra posibilidad es que estas alteraciones
Esto podría explicar que, en estudios de fuentes de corriente, se
del control gabérgico se relacionen con la reorganización del
La alteración más típica del hipocampo
epiléptico es la esclerosis, caracterizada porla reinervación de los gránulos del giro denta-do por las fibras musgosas y la muerte neuro-nal en el hilus, zonas de CA3 y CA1 y las ca-pas superficiales de la corteza entorrinal (Figs. 1c y 1d) [55]. La reinervación o sprouting pa-rece no guardar relación con la epileptogéne-sis, por lo que se ha sugerido que es una con-secuencia –más que una causa– de las crisis. En el kindling, por ejemplo, las crisis puedenllegar a generalizarse mucho antes de que sedetecte esclerosis del hipocampo [56]. Esto
no parece ser exclusivo de este modelo, pues-to que en ratas tratadas con pilocarpina o kai-nato, el bloqueo de la reinervación con el in-hibidor de síntesis proteica cicloheximida no
A pesar de esta evidencia, se piensa que la
reinervación por las fibras musgosas constitu-ye un mecanismo de amplificación de las des-cargas. Registros in vitro han demostrado queestos circuitos aberrantes glutamatérgicos sonfuncionales y favorecen la sincronizaciónneuronal [58]. Se han detectado respuestas en
descargas en el giro dentado y mayor activa-
Figura 1. Estudios fisiopatológicos en modelos animales de ELT. a) Descarga registrada in vitro
ción de las dendritas de los gránulos [59]. Los
en rodajas de hipocampo de ratas tratadas con litio-pilocarpina; nótese la presencia de ritmos
circuitos reinervados son de naturaleza gluta-
ultrarrápidos. b) Espectro de potencia de registros de campo en rodajas de hipocampo de ratas
matérgica, y estudios moleculares han de-
control y epilépticas (modelo litio-pilocarpina); nótense los componentes de alta frecuenciacorrespondientes a los ritmos ultrarrápidos presentes en el caso epiléptico. c) Tinción de Nissl
mostrado que existen incrementos en las sub-
en hipocampo de ratas tratadas con litio-pilocarpina que muestra la muerte en la región del
hilus asociada con la esclerosis del hipocampo (flecha); barra de calibración: 250 µm. d) Tinción
asociado al sprouting, lo que sugiere una res-
de Timm en hipocampo de ratas tratadas con litio-pilocarpina que muestra la reinervación porlas fibras musgosas característica de la esclerosis del hipocampo (flecha); barra de calibración:
puesta incrementada a través de estos recep-
tores [60]. Igualmente, se han detectado alte-raciones en la región CA3, en particular de lasubunidad GluR2 del receptor AMPA. Puesto
sistema inhibidor detectada en el hipocampo y la corteza ento-
que un decremento de esta subunidad origina receptores AMPA
rrinal en la fase crónica (véase el siguiente apartado).
con mayor permeabilidad al Ca2+, se ha propuesto que éste
En resumen, la contribución de la neocorteza parece ser fun-
constituya un importante mecanismo de epileptogénesis (hipó-
damental en las fases iniciales del desarrollo de la epilepsia.
tesis GluR2) [61]. De hecho, ratones modificados genéticamen-
Alteraciones del sistema glutamatérgico, unido a la gran excita-
te para expresar una forma no editada de la subunidad GluR2
bilidad del circuito local en las cortezas piriforme y perirrinal,
muestran crisis tempranas y alta mortalidad a las tres semanas
parecen ser en gran medida responsables de la tendencia de es-
tas estructuras a generar posdescargas que facilitan el recluta-
Otra de las alteraciones asociadas a la reinervación por las
miento del hipocampo y la corteza entorrinal en lazos de activi-
fibras musgosas guarda relación con los receptores glutamatér-
gicos de kainato, que desempeñan un papel fundamental en elcontrol presináptico y postsináptico de la excitabilidad. La ex-
Alteraciones en el hipocampo
presión de la subunidad KA1 está incrementada de manera per-
Tanto desde el punto electrofisiológico como anatomopatológi-
manente en la región CA3, pero no en el giro dentado de ratas
co, el hipocampo es la estructura más estudiada en la ELT. El
sometidas a kindling [63]. Otros autores han demostrado un
EEG de animales epilépticos muestra claros signos de actividad
incremento transitorio de la subunidad KA2 [64]. Un estudio de
interictal, aunque éstos se desarrollan antes en otras zonas, co-
ganancia de función, en el que se transfectó el ARN para GluR6
mo las cortezas perirrinal y piriforme [53]. El mapeo funcional
en rodajas de hipocampo en cultivo, mostró un incremento de
visualizando consumo de 2-deoxiglucosa y expresión de c-fos
la excitabilidad y mayor propensión a la generación de descar-
demuestra que el hipocampo es de las últimas estructuras en ac-
gas epileptiformes [65]. Curiosamente, se ha encontrado una
tivarse en el kindling de la amígdala y la corteza [54]. Sin em-
mayor susceptibilidad a la epilepsia en ratas que sobreexpresan
bargo, la esclerosis del hipocampo, principal patología en la
ELT en humanos, se reproduce en los modelos animales, y las
No sólo los receptores ionotrópicos de glutamato muestran
alteraciones fisiológicas son evidentes en la fase crónica (Figs.
niveles alterados de expresión en el hipocampo epiléptico. En
1a y 1b). Esto sugiere un papel importante del hipocampo en la
ratas sometidas al modelo de estado epiléptico o kindling, se ha
descrito una menor sensibilidad de los receptores metabotrópi-
cos de los grupos II y III a los agonistas AP4, PPG y CCG [67].
un potencial postsináptico inhibitorio con componentes rápidos
Como estos receptores presumiblemente actúan como autorre-
y lentos mediados por los receptores GABA y GABA , respec-
ceptores presinápticos, una alteración de su funcionalidad afec-
tivamente [86,90-92]. En algunos trabajos se ha mostrado que
ta sensiblemente la regulación de la liberación de glutamato. De
cerca del 80% de las células registradas en regiones próximas a
hecho, en la región CA3 se han mostrado incrementos en la li-
zonas epileptógenas generan despolarizaciones intensas sin inhi-
beración de glutamato dependiente de Ca2+ [68].
bición posterior, lo que sugiere una alta excitabilidad en el cir-
Otra de las alteraciones más importantes en el hipocampo
cuito y cierto grado de desinhibición [93,94]. Estas despolariza-
epiléptico afecta a la heterogénea población de células gabérgi-
ciones, que se acompañan de ráfagas de potenciales de acción,
cas y, por consiguiente, a la inhibición. Se ha descrito que la
están mediadas por receptores NMDA fundamentalmente [86].
desconexión glutamatérgica de interneuronas del hilus origina
Aunque la fisiología estudiada en humanos reproduce muy
una desinhibición del giro dentado y la región CA3 que facilita
fielmente la encontrada en modelos animales, hay un aspecto en
la generación de descargas [69]. Sin embargo, otros trabajos han
el que no existe paralelismo, y es la presencia de potenciales in-
mostrado que el decremento de la inhibición se recupera pro-
hibitorios espontáneos generalizados en el tejido humano [95-
gresivamente en paralelo a la reinervación por las fibras musgo-
97]. Estos potenciales están ampliamente distribuidos por la
sas [70,71]. Consecuentemente, en el giro dentado se registra un
corteza, pueden registrarse de forma sincrónica en regiones cla-
incremento en la depresión por pares de pulsos, lo que sugiere
ramente separadas y poseen una morfología similar a las puntas
un aumento de la inhibición que es dependiente de la activación
u ondas agudas observadas clínicamente en el EEG [98]. El
de receptores NMDA [72]. En ratas tratadas con pilocarpina o
estudio intracelular ha demostrado que estos eventos revierten
kainato, también se registra un decremento transitorio de la
en torno a –70 mV y parecen estar mediados por una conduc-
inhibición que se recupera progresivamente conforme avanza el
tancia de Cl– vinculada a receptores GABA [97,98]. No obs-
proceso de epileptogénesis [73]. En contraste, en la región CA1,
tante, es importante destacar que gran parte de estos trabajos se
la inhibición por pares está disminuida, y concretamente, en el
han realizado sobre la corteza temporal lateral en pacientes con
modelo de kindling, se ha visto que esta disminución es progre-
ELT asumiendo que se trata de una región libre de patología y
siva durante el protocolo de estimulación [74,75]. Usando fuen-
que, por tanto, sirve como modelo para el estudio fisiológico
tes de corrientes se ha demostrado que, en el modelo de estado
[78]. Sin embargo, esta presunción está muy lejos de ser verdad,
epiléptico, el potencial postsináptico excitatorio registrado en
como lo prueba el hecho de que más de un 75% de los pacientes
radiado se incrementa por pulsos pareados, lo que sugiere un
intervenidos de ELT presentan actividad interictal significativa
incremento en la excitabilidad dendrítica provocado por la des-
en la región lateral durante los estudios de electrocorticografía
(ECoG) [88,99]. En cualquier caso, conviene destacar que unaactividad similar se ha descrito en registros in vitro del subículode pacientes intervenidos de ELT que mostraban esclerosis del
ESTUDIOS EN HUMANOS
Los datos fisiopatológicos de tejido de pacientes con epilepsia
El estudio de la actividad interictal en tejido humano epilép-
son, por razones obvias, claramente más limitados que los obte-
tico in vitro ha permitido abordar los posibles mecanismos res-
nidos a partir de modelos animales. Sin embargo, en los últimos
ponsables de la hiperexcitabilidad. En la mayoría de estos traba-
años, gracias a la mejora técnica en determinados métodos diag-
jos, no se ha registrado actividad interictal espontánea en me-
nósticos y a la disposición de tejido procedente de pacientes
dios normales (véase, sin embargo, Cohen et al [89]) y se han
intervenidos de epilepsia, el avance ha sido espectacular [78].
utilizado diferentes agentes farmacológicos para inducirla y es-tudiar las alteraciones en los circuitos. Así, se ha podido com-
Fisiopatología de la región neocortical
probar que la inhibición de los receptores GABA genera des-
El estudio de las muestras de tejido obtenido a partir de pacien-
cargas paroxísticas de tipo epileptógeno de morfología similar a
tes intervenidos por epilepsia focal farmacorresistente tiene una
la estudiada en animales [86,100,101] y en la que participan los
extraordinaria importancia y ha permitido desvelar propiedades
receptores NMDA de glutamato [102,103]. En cambio, en las
electrofisiológicas básicas del tejido humano, tanto epileptóge-
descargas inducidas por 4AP, la contribución de conductancias
no como no epileptógeno. Básicamente, la electrofisiología del
al bicarbonato mediadas por los receptores de GABA resulta
tejido neocortical humano es similar a la estudiada en modelos
fundamental [104-106]. El bloqueo de los receptores GABAA
animales. En neuronas neocorticales humanas se han descrito
en estas condiciones prolonga las descargas, las cuales no son
conductancias de Na+, K+ y Ca2+ con características similares a
eliminadas por antagonistas de receptores glutamatérgicos [107].
las estudiadas en animales [79-85]. En respuesta a la inyección
Se ha sugerido que las descargas interictales en el tejido huma-
intracelular de corriente, estas células responden con series de
no precisan no sólo de la participación de conductancias media-
potenciales de acción en dos rangos lineales diferentes [86,87],
das por receptores de tipo GABA , sino de la inhibición de los
presentando además adaptación. Por su parte, las interneuronas
receptores presinápticos de tipo GABA , que presuntamente se
registradas en las zonas que muestran actividad de tipo epilepti-
localizan en interneuronas [107]. Otro fenómeno que presumi-
forme in situ, exhiben una electrofisiología similar a la descrita
blemente contribuye a la sincronización de extensas regiones
en la corteza de rata, consistente en el disparo rápido de poten-
corticales durante las espigas interictales es la elevación de la
ciales de acción en respuesta a pulsos supraumbrales [88]. En el
concentración extracelular de K+, que podría despolarizar inter-
caso del subículo, se ha descrito además la presencia de neuro-
neuronas vecinas, aumentando de este modo la extensión de la
nas con capacidad de disparo en ráfagas [89].
Las células neocorticales suelen responder ortodrómicamen-
Con relación a las alteraciones funcionales y estructurales
te con un potencial postsináptico excitatorio, mediado de forma
es importante destacar que los cambios encontrados en las zo-
fundamental por receptores no NMDA de glutamato, seguido de
nas corticales asociadas a la esclerosis del hipocampo en la ELT
se han estudiado menos que aquellos relacionados con síndro-
de control de la excitabilidad de un gran número de células pi-
mes que cursan con epilepsia, como la displasia cortical [109].
No obstante, en aquellas regiones temporales que muestran ac-
La electrofisiología de la región temporal mesial en la ELT
tividad interictal en la ECoG intraoperatoria, se ha encontrado
es similar a la observada para la región neocortical y a la hallada
un decremento de interneuronas gabérgicas, fundamentalmente
en modelos animales [93,125,126]. Llama la atención el hecho
de las células en cesto positivas a parvoalbúmina [110]. Igual-
de que, en general, las propiedades electrofisiológicas de las neu-
mente, se han detectado parches en la inmunorreactividad con-
ronas de las regiones epileptógenas no se modifiquen, de modo
tra parvoalbúmina y calbindina, lo cual sugiere que las altera-
que variables como la resistencia de entrada, el potencial de re-
ciones son microanatómicas [111]. Estudios inmunocitoquími-
poso o las propiedades de descarga no cambian con respecto a
cos contra diferentes subunidades de los receptores glutamatér-
neuronas de regiones no epileptógenas [127]. Sin embargo, se
gicos han mostrado un incremento en la expresión de la subuni-
han observado cambios importantes en la expresión de ARNm
dad NMDAR2 en neuronas cercanas a la zona irritativa [112],
para los canales catiónicos HCN activados por hiperpolarización
así como alteraciones en la expresión de NMDAR1, GluR2 y
y modulados por nucleótidos cíclicos en las células del giro
GluR5 [113]. Asimismo, se ha informado de defectos en la efi-
dentado [128], aunque este efecto no se ha estudiado electrofi-
ciencia de edición de las subunidades GluR5 y GluR6 en la cor-
siológicamente. Además, se ha comprobado la disminución de
teza temporal de pacientes con ELT [114].
amplitud de la posthiperpolarización en aquellas regiones con
También se han descrito alteraciones importantes en la ex-
capacidad de generar actividad epiléptica espontánea [127].
presión génica del neocórtex temporal en la ELT. En efecto, re-
Los cambios electrofisiológicos más significativos encon-
cientemente, utilizando la técnica de microarrays se ha mues-
trados en rodajas de hipocampo se han observado en la activi-
treado la expresión de gran número de genes en tejido humano
dad sináptica. De hecho, se ha podido comprobar que los gránu-
[115]. Así, sobre una muestra de 14.500 genes, se ha podido de-
los del giro dentado son capaces de generar potenciales sinápti-
mostrar que existe una expresión diferencial de genes en distin-
cos dependientes de NMDA prolongados, mientras que la con-
tas áreas corticales, dependiendo de que muestren o no activi-
ductancia de receptores AMPA es normal [129]. En este mismo
dad epileptógena in situ [116]. En este sentido, en las regiones
sentido, se ha descrito el aumento en los niveles de expresión de
definidas como epileptógenas o irritativas se observó una dis-
los receptores de NMDA y AMPA, que en el caso de los prime-
minución en la expresión diferencial de 30 genes, mientras que
ros, se correlaciona con incrementos en la expresión de las sub-
pudo verse un incremento en la expresión de otros 46 genes.
unidades NR2A y NR2B, tanto en células del giro dentado co-
Entre los genes cuya expresión se encuentra disminuida se ha-
mo en neuronas de CA1 [130-132]. En pacientes con esclerosis
llan genes de señalización, genes que codifican diversas señales
se ha comprobado, también, que hay una menor expresión de la
gabérgicas y genes expresados por oligodendrocitos, mientras
subunidad NR2A en las áreas CA2/3. Además, similar a lo des-
que en esta misma región se observó un incremento muy im-
crito en animales, se ha informado de una pérdida de función de
portante del gen de transferrina, cuya significación aún resulta
los receptores metabotrópicos que actúan presinápticamente so-
bre el control de la liberación de glutamato [133].
Con relación a la inhibición, varios trabajos han mostrado
Fisiopatología de la región mesial
que el número de receptores o subunidades de tipo GABA está
La lesión anatomopatológica más frecuente de la ELT es la es-
reducido en el hipocampo esclerótico [134-136]. Utilizando téc-
clerosis mesial, que representa hasta el 70% de las lesiones ob-
nicas inmunohistoquímicas se ha informado de la sobrerregula-
tenidas a partir de series quirúrgicas [109] y que, como se ha
ción de la subunidad 2 del receptor de GABA en el soma y en
mencionado, se caracteriza por una pérdida selectiva de neuro-
las dendritas apicales, y de un marcaje reducido en las dendritas
nas en las capas superficiales del córtex entorrinal, especial-
basales [137]. Igualmente, se ha observado reorganización de
mente en la porción medial, en el hilus del dentado y en las áre-
los terminales gabérgicos de las células en candelabro en el giro
as CA1 y CA3 del hipocampo [117]. Otros hallazgos, como la
dentado, en la formación hipocámpica y en el subículo [119].
dispersión de los gránulos y la pérdida selectiva de células hila-
En este último caso, se ha descrito la reinervación de los axones
res portadoras de somatostatina y neuropéptido Y [118], tam-
de las células en candelabro que forman agregados hipertrófi-
bién contribuyen a la caracterización de esta entidad. Sin em-
cos. No está claro aún si esto representa un caso de incremento
bargo, las modificaciones histopatológicas están lejos de ser ho-
de la inhibición, puesto que se ha visto que en estas células tie-
mogéneas como para identificar un conjunto unívoco de lesio-
nen lugar alteraciones en la expresión de los cotransportadores
nes hipocampales responsables de la epilepsia [119]. En este
de Cl– –NKCC1 y KCC2–, responsables de la homeostasis in-
sentido, cobra fuerza la idea de que, como ha ocurrido en los es-
tudios sobre animales, la esclerosis del hipocampo es una pato-
Además de posibles efectos en la inervación excitatoria so-
logía secundaria al proceso epileptógeno.
bre interneuronas gabérgicas, se ha observado que la funcionali-
Además de la pérdida neuronal, son dos los aspectos anato-
dad de éstas permanece alterada, presentando una recuperación
mopatológicos más relevantes de la esclerosis en humanos. Por
más lenta ante trenes de alta frecuencia que el hipocampo nor-
un lado, se observa una intensa proliferación de axones abe-
mal [139]. Registros electrofisiológicos en rodajas de hipocam-
rrantes desde los gránulos [120-122], similar a lo previamente
po in vitro han demostrado que la región del subículo, con una
descrito en modelos animales. Consistente con esta reinerva-
amplia variabilidad topográfica, genera eventos sincrónicos es-
ción se ha encontrado un incremento significativo de la den-
pontáneos mediados por los receptores de tipo GABA [89] y
sidad de espinas dendríticas en las células del giro dentado
de morfología similar a las descargas interictales. Durante estos
[123]. Por otro lado, existe una pérdida selectiva de células
potenciales de campo, aumenta el disparo de potenciales de ac-
candelabro en la región de transición entre CA1 y el subículo,
ción en interneuronas, mientras que en las células piramidales
que constituyen interneuronas inhibitorias con gran capacidad
se detectan tanto eventos hiperpolarizantes como despolarizan-
tes. El efecto despolarizante parece de-berse a un aumento anómalo de la con-
centración intracelular de Cl–, presumi-blemente debido a alteraciones de los co-
transportadores KCC2 y NKCC1, querevertiría la dirección de la corrienteinducida por la apertura de los canales
sión disminuida del cotransportador KCC2es una consecuencia de la desaferentiza-
ción de CA1 observada en la ELT [140,141], aunque también hay resultados si-milares en tejido no esclerótico [127].
ciones en la región mesial, cuya interpre-tación es aún incierta y se encuentran eninvestigación. Es el caso del incrementoen la expresión de los receptores Y2 y ladisminución de los receptores Y1 –am-bos del neuropéptido Y– en el hipocam-po de pacientes con ELT [142]. Como elneuropéptido Y regula la actividad de in-terneuronas y el flujo sináptico de Ca2+[143], las alteraciones descritas previa-mente podrían tener repercusión sobre laexcitabilidad del hipocampo. Otro inte-
resante mecanismo que se está estudian-do activamente es la participación de lasuniones eléctricas en el control de la ex-citabilidad. Se ha observado una eleva-ción de ARNm de conexina 43 en tejidoepiléptico [144,145], lo que indicaría queun incremento de la presencia de estaproteína podría modificar las propieda-des de sincronización del tejido. Figura 2. Estudios fisiopatológicos en pacientes con ELT. a) Tipos de distribución de la actividad in- ESTUDIOS INVASIVOS
terictal en ELT registrada mediante electrodos de foramen oval: a1) Distribución de frecuencia muy
EN PACIENTES CON ELT:
dispersa de las fuentes de voltaje por gran parte de la región mesial en un paciente con ELT; a2) Dis-tribución de frecuencia localizada de otro paciente, en la que las fuentes de voltaje se localizaron
PERSPECTIVAS DE FUTURO
con menos de 1 cm de dispersión. b) Registros extracelulares realizados in situ sobre la corteza tem-
La necesidad de realizar estudios invasi-
poral lateral de un paciente durante la cirugía: b1) Respuesta neuronal durante la aplicación intra-venosa de etomidato (0,1 mg/kg); b2) Detalle de uno de los potenciales de acción registrados en b1.
vos al objeto de identificar el foco epi-
c) Características fisiopatológicas de las regiones temporal lateral y mesial registradas durante la
ECoG y analizadas mediante técnicas no lineales: c1) Se muestra la matriz de correlación de la acti-
tidad de información sobre la fisiopato-
vidad registrada en mantas de 20 canales EEG colocadas en la corteza temporal y tiras de entre 4 y8 canales colocadas en la cara mesial; c2) Se muestra la representación en la corteza temporal
logía de la epilepsia. Estudios con elec-
(manta) y mesial (tira) de la dinámica de correlación y sincronización de fase.
trodos profundos han mostrado una ma-yor resolución para detectar las zonas deorigen de la actividad interictal e ictal en la ELT, localizándose
bar que existen pacientes en los que la actividad interictal está
generalmente en las zonas de la región hipocampal y parahipo-
ampliamente distribuida por la región mesial, mientras que en
campal [146,147]. La utilización de estas técnicas permite, por
otros está mucho más limitada (Figs. 2a1 y 2a2). Este mismo
tanto, acceder electrofisiológicamente a las zonas de generación
tipo de abordaje nos ha permitido estudiar la relación entre la
de la actividad paroxística, lo que resulta de gran utilidad para
región irritativa (responsable de las manifestaciones intericta-
abordar diferentes aspectos de los mecanismos básicos in situ.
les) y la región de comienzo ictal, además de conocer la rela-
Por ello, no podíamos terminar sin efectuar un breve repaso de
ción entre la atrofia y la esclerosis observada en la resonancia
las posibilidades presentes y futuras de este tipo de estudios.
magnética y la topografía de la actividad bioeléctrica.
El estudio con electrodos de foramen oval (EFO) en pacien-
Otra técnica de gran importancia es la utilización de microe-
tes con ELT se introdujo como una alternativa semiinvasiva pa-
lectrodos profundos para el estudio de células in situ. Emplean-
ra caracterizar prequirúrgicamente la fisiopatología de la región
do ramilletes de microelectrodos de unos 50 μm de diámetro es
mesial [148-150]. Recientemente, hemos aplicado la aproxima-
posible registrar la actividad aislada de neuronas próximas y
ción monopolar al estudio de la actividad interictal e ictal regis-
relacionarla con el EEG local (Figs. 2b1 y 2b2) [152,153]. La
trada con EFO [151]. Esta cuantificación nos permitió compro-
utilización de nuevos sistemas de amplificación con multicana-
les y el desarrollo de herramientas computacionales de análisis
nar importantes pistas sobre la dinámica de la actividad inter-
ha permitido ampliar el estudio a toda la banda de frecuencias,
ictal y el estudio de la anticipación de crisis [163,164]. Una
tradicionalmente limitado en el EEG [154]. De este modo se ha
aproximación es el estudio de las señales registradas mediante
podido mostrar que las neuronas de la región epileptógena tie-
ECoG intraoperatoria en la ELT. Este abordaje permite perfilar
nen una mayor frecuencia de descarga y mayor propensión a la
las zonas en la corteza temporal lateral que generan actividad
agrupación de su actividad en forma de ráfagas [155], y se ca-
sincronizada y caracterizar su dinámica espaciotemporal (Figs.
racterizan por la presencia de descargas sincrónicas de muy alta
2c1 y 2c2). Actualmente existe cada vez más consenso en que la
frecuencia (200-500 Hz), denominadas ritmos ultrarrápidos o
aplicación de métodos de análisis no lineales proporciona mu-
fast ripples [147]. Algunos autores consideran que los ritmos
cha más información sobre los diferentes procesos básicos res-
ultrarrápidos constituyen potenciales inhibidores sincronizados
ponsables de las crisis y su utilidad diagnóstica se ha sugerido
sobre los somas de las neuronas principales [156], aunque otros
piensan que estas oscilaciones estarían originadas por reverbe-raciones a través de uniones eléctricas [157].
La utilización de electrodos profundos para estudiar poten-
CONCLUSIONES
ciales de campo in situ en pacientes con ELT ha permitido tam-
En resumen, los estudios sobre la fisiopatología de la ELT en
bién analizar la plasticidad sináptica a corto plazo en regiones
animales y humanos sugieren una gran heterogeneidad en las
escleróticas. Estos trabajos han mostrado que la depresión de
alteraciones responsables de la hiperexcitabilidad. Aunque
pulsos pareados está incrementada en la vía perforante y dismi-
resulta difícil correlacionar cambios específicos, existe una base
nuida en las vías de asociación intrínsecas del hipocampo escle-
común expresada en forma de remodelación de la excitación y
rótico [158]. Esta inhibición excesiva en la principal vía de
la inhibición que deriva en hiperexcitabilidad y sustenta la ge-
entrada al hipocampo se ha interpretado más como un mecanis-
neración de descargas paroxísticas. En cualquier caso, es im-
mo de protección que como un proceso desencadenante de la
portante no perder de vista que una crisis epiléptica es el re-
patología, en tanto que la disminución de la inhibición en las
sultado clínico de una descarga brusca –anormal por su intensi-
vías de asociación podría contribuir al aumento de sincroniza-
dad e hipersincronía– de un conjunto de neuronas del cerebro, y
ción responsable de la actividad epileptógena. En este mismo
por lo tanto, cualquier alteración que favorezca la hipersincroni-
sentido, se ha observado que la potenciación a largo plazo está
zación y la excitabilidad es susceptible de originar descargas
disminuida en estos pacientes, lo que podría explicar los pobres
resultados en determinadas tareas neuropsicológicas que requie-
Es mucho lo que se sabe sobre diversos aspectos fisiopato-
ren la participación de la memoria [159].
lógicos de la ELT, tanto en modelos animales como en huma-
Para finalizar, mencionar muy brevemente una última apro-
nos; sin embargo, es tanto o más lo que todavía se desconoce
ximación al estudio de la fisiopatología de la epilepsia, como es
acerca de las causas desencadenantes, los procesos fisiopatoló-
el desarrollo y la aplicación de herramientas matemáticas de
gicos reales que explican la génesis de las crisis, su clínica y
análisis no lineal a diferentes tipos de registros electrofisiológi-
evolución. Todas estas cuestiones se relacionan íntimamente
cos. A lo largo de estos años se han aplicado diferentes medidas
con los mecanismos fisiopatológicos y, por ello, un conoci-
no lineales que permiten la caracterización de las propiedades
miento lo más completo posible de éstos puede ser de gran im-
dinámicas de las series de tiempo, como los exponentes de Lya-
portancia no sólo desde el punto de vista neurocientífico, sino
punov [160], la dimensión de correlación [161] y las medidas
especialmente desde el punto de vista médico, permitiendo ofre-
de interdependencia [162], por citar sólo algunas. La aplicación
cer nuevas opciones terapéuticas a los pacientes aquejados por
a registros preoperatorios e intraoperatorios parece proporcio-
1. Berger H. Uber das Electrenkephalogram des Menschen [On the EEG
11. Purpura DP. Nature of electrical potentials and synaptic organizations
in humans]. Arch Psychiatr Nervenkr 1929; 87: 527-570.
in cerebral and cerebellar cortex. Int Rev Neurobiol 1959; 1: 47-163.
2. Gibbs FA, Davis H, Lennox WG. The EEG in epilepsy and in the
12. Kandel ER, Spencer WA. The pyramidal cell during hippocampal
impaired states of consciousness. Arch Neurol Psychiatry 1935; 34: 1133.
3. Gibbs FA, Lennox WG, Gibbs EL. The electroencephalogram in diag-
13. Kandel ER, Spencer WA. Electrophysiology of hippocampal neurons.
nosis and in localization of epileptic seizures. Arch Neurol Psychiatry
I. Sequential invasion and synaptic organization. J Neurophysiol 1961;
4. Jasper HH. Localized analyses of the function of the human brain by
14. Kandel ER, Spencer WA. Electrophysiology of hippocampal neurons. II.
the electro-encephalogram. Arch Neurol Psychiatry 1936; 36: 1131.
After-potentials and repetitive firing. J Neurophysiol 1961; 24: 243-59.
5. Thakor NV, Shanbao T. Advances in quantitative electroencephalo-
15. Goldensohn ES, Purpura DP. Intracellular potentials of cortical neu-
gram analysis methods. Ann Rev Biomed Eng 2004; 6: 453-95.
rons during focal epileptogenic discharges. Science 1963; 139: 840-2.
6. Alarcon G, Binnie CD, Elwes RDC, Polkey. CE Power spectrum and
16. Matsumoto H, Amjone-Marsane C. Cortical cellular phenomena in expe-
intracranial EEG patterns at seizure onset in partial epilepsy. Electro-
rimental epilepsy: interictal manifestations. Exp Neurol 1964; 9: 286-304.
encephalogr Clin Neurophysiol 1995; 94: 326-37.
17. Matsumoto H, Amjone-Marsane C. Cortical cellular phenomena in ex-
7. Urrestarazu E, Iriarte J. Análisis matemático en el estudio de señales
perimental epilepsy: ictal manifestations. Exp Neurol 1964; 9: 305-26.
electroencefalográficas. Rev Neurol 2005; 41: 423-34.
18. Creutzfeldt O, Watanabe S, Lux HD. Relations between EEG phenom-
8. Goldensohn ES. Historical perspectives. In Engel J Jr, Pedley TA, eds.
ena and potentials of single cortical cells. II. Spontaneous and convul-
Epilepsy: a comprehensive textbook. Vol. I. Philadelphia: Lippincott
soid activity. Electroencephalogr Clin Neurophysiol 1966; 20: 19-37.
Williams & Wilkins; 1997. p. 15-39.
19. Prince DA. The depolarization shift in ‘epileptic’ neurons. Exp Neurol
9. Renshaw B, Forbes A, Morisno BR. Activity of isocortex and hip-
pocampus: electrical studies with microelectrodes. J Neurophysiol 1940;
20. Goldensohn ES, Zablow L, Salazar AM. The penicillin focus. I. Distri-
bution of potential at the cortical surface. Electroencephalogr Clin Neuro-
10. Li CH, Jasper HH. Microelectrode studies of electrical activity of the
cerebral cortex in the cat. J Physiol (Lond) 1953; 121: 117-40.
21. Goldensohn ES, Zablow L, Stein B. Interrelationships of form and la-
tency of spike discharge from small areas of human cortex. Electroen-
48. Namba T, Morimoto K, Sato K, Yamada N; Kuroda S. Antiepilepto-
cephalogr Clin Neurophysiol 1970; 29: 321-2.
genic and anticonvulsant effects of NBQX, a selective AMPA receptor
22. Goddard GV. Development of epileptic seizures through brain stimula-
antagonist, in the rat kindling model of epilepsy. Brain Res 1994; 638:
tion at low intensity. Nature 1967; 214: 1020-1.
23. Racine RJ. Modification of seizure activity by electrical stimulation. I.
49. Prince HK, Conn PJ, Blackstone CD, Huganir RL, Levey AI. Down-
After-discharge threshold. Electroencephalogr Clin Neurophysiol 1972;
regulation of AMPA receptor subunit GluR2 in amygdaloid kindling. J
24. Racine RJ. Modification of seizure activity by electrical stimulation. II.
50. Racine RJ, Moore KA, C Evans. Kindling-induced potentiation in the
Motor seizure. Electroencephalogr Clin Neurophysiol 1972; 32: 281-94.
piriform cortex. Brain Res 1991; 556: 218-25.
25. Pinel JP, Rovner LI. Experimental epileptogenesis: kindling-induced
51. Wilson CL, Khan SU, Engel JJr, Isokawa M, Babb TL, Behnke EJ.
epilepsy in rats. Exp Neurol 1978; 58: 190-202.
Paired pulse suppression and facilitation in human epileptogenic hip-
26. Morimoto K, Sato H, Sato K, Sato S, Yamada N. BW1003C87, pheny-
pocampal formation.Epilepsy Res 1998; 31: 211-30.
toin and carbamazepine elevate seizure threshold in the rat amygdala-
52. Lehmann H, Ebert U, Loscher W. Amygdala-kindling induces a lasting
kindling model of epilepsy. Eur J Pharmacol 1997; 339: 11-5.
reduction of GABA-immunoreactive neurons in a discrete area of the
27. Ben-Ari Y. Limbic seizure and brain damage produced by kainic acid:
ipsilateral piriform cortex. Synapse 1998; 29: 299-309.
mechanisms and relevance to human temporal lobe epilepsy. Neuro-
53. Kairiss EW, Racine RJ, Smith GK. The development of the interictal
spike during kindling in the rat. Brain Res 1984; 322: 101-10.
28. Turski WA, Cavalheiro EA, Schwarz M, Czuczwar SJ, Kleinrok Z,
54. Sato T, Yamada N, Morimoto K, Uemura S, Kuroda S. A behavioral
Tursk L. Limbic seizures produced by pilocarpine in rats: behavioural,
and immunohistochemical study on the development of perirhinal cor-
electroencephalographic and neuropathological study. Behav Brain Res
tical kindling: a comparison with other types of limbic kindling. Brain
29. Tremblay E, Nitecka L, Berger ML, Ben-Ari Y. Maturation of kainic
55. Nadler JV, Perr BW, Cotman CW. Selective reinnervation of hippo-
acid seizure-brain damage syndrome in the rat. I. Clinical, electro-
campal area CA1 and the fascia dentata after destruction of CA3-CA4
graphic and metabolic observations. Neuroscience 1984; 13: 1051-72.
afferents with kainic acid. Brain Res 1980; 182: 1-9.
30. Lothman EW, Hatlelid JM, Zorumski CF. Functional mapping of lim-
56. Ebert U, Loscher W. Differences in mossy fibre sprouting during con-
bic seizures originating in the hippocampus: a combined 2-deoxyglu-
ventional and rapid amygdala kindling of the rat. Neurosci Lett 1995;
cose and electrophysiologic study. Brain Res 1985; 360: 92-100.
31. Saji M, Kobayashi S, Ohno K, Sekino Y. Interruption of supramammil-
57. Longo BM, Mello LE. Blockade of pilocarpine- or kainate-induced
lohippocampal afferents prevents the genesis and spread of limbic
mossy fiber sprouting by cycloheximide does not prevent subsequent
seizures in the hippocampus via a disinhibition mechanism. Neuro-
epileptogenesis in rats. Neurosci Lett 1997; 226: 163-6.
58. Scharfman HE, Sollas AL, Berge RE, Goodman JH. Electrophysiolog-
32. Racine RJ, Mosher M, Kairiss EW. The role of the pyriform cortex in
ical evidence of monosynaptic excitatory transmission between gran-
the generation of interictal spikes in the kindled preparation. Brain Res
ule cells after seizure-induced mossy fiber sprouting. J Neurophysiol
33. McIntyre DC, Plant JR. Long-lasting changes in the origin of sponta-
59. Otsu Y, Maru E, Ohata H, Takashima I, Kajiwara R, Iijima T. Optical
neous discharges from amygdala-kindled rats: piriform vs. perirhinal
recording study of granule cell activities in the hippocampal dentate
cortex in vitro. Brain Res 1993; 624: 268-76.
gyrus of kainate-treated rats. J Neurophysiol 2000; 83: 2421-30.
34. McIntyre DC, Wong RK. Cellular and synaptic properties of amyg-
60. Mikuni N, Babb TL, Wylie C; Ying Z. NMDAR1 receptor proteins and
dala-kindled piriform cortex in vitro. J Neurophysiol 1986; 55: 1295-307.
mossy fibers in the fascia dentata during rat kainate hippocampal
35. Gernert M, Bloms-Funke P, Ebert U, Loscher W. Kindling causes per-
epileptogenesis. Exp Neurol 2000; 163: 271-7.
sistent in vivo changes in firing rates and glutamate sensitivity of cen-
61. Friedman LK, J Veliskova. GluR2 hippocampal knockdown reveals
tral piriform cortex neurons in rats. Neuroscience 2000; 99: 217-27.
developmental regulation of epileptogenicity and neurodegeneration.
36. Haberly LB, Sutula TP. Neuronal processes that underlie expression of
Brain Res Mol Brain Res 1980; 61: 224-31.
kindled epileptiform events in the piriform cortex in vivo. J Neurosci
62. Brusa R, Zimmermann F, Koh DS, Feldmeyer D, Gass P, Seeburg PH,
et al. Early-onset epilepsy and postnatal lethality associated with an
37. Lothman EW, Bertram EH, Stringer JL. Functional anatomy of hip-
editing-deficient GluR-B allele in mice. Science 1995; 270: 1677-80.
pocampal seizures. Prog Neurobiol 1991; 37: 1-82.
63. Hikiji M, Tomita H, Ono M, Fujiwara Y, Akiyama K. Increase of kai-
38. Kelly ME, McIntyre DC. Perirhinal cortex involvement in limbic kin-
nate receptor mRNA in the hippocampal CA3 of amygdala-kindled
dled seizures. Epilepsy Res 1996; 26: 233-43.
rats detected by in situ hybridization. Life Sci 1993; 53: 857-64.
39. Morimoto K, Dragunow M, Goddard GV. Deep prepyriform cortex
64. Kamphuis W, Hendriksen H, Diegenbach PC, Lopes da Silva FH. N-
kindling and its relation to amygdala kindling in the rat. Exp Neurol
methyl-D-aspartate and kainate receptor gene expression in hippocam-
pal pyramidal and granular neurons in the kindling model of epilepto-
40. Bengzon J, Okabe S, Lindvall O, McKay RD. Suppression of epilepto-
genesis. Neuroscience 1995; 67: 551-9.
genesis by modification of N-methyl-D-aspartate receptor subunit com-
65. Bergold PJ, Casaccia-Bonnefil P, Zeng XL, Federoff. HJ Transsynaptic
position. Eur J Neurosci 1999; 11: 916-22.
neuronal loss induced in hippocampal slice cultures by a herpes sim-
41. Kikuchi S, Iwasa H, Sato T. Lasting changes in NMDAR1 mRNA lev-
plex virus vector expressing the GluR6 subunit of the kainate receptor.
el in various regions of cerebral cortex in epileptogenesis of amyg-
daloid-kindled rat. Psychiatry Clin Neurosci 2000; 54: 573-7.
66. Telfeian AE, Federoff HJ, Leone P, During MJ, Williamson A. Overex-
42. Kraus JE, McNamara JO. Measurement of NMDA receptor protein
pression of GluR6 in rat hippocampus produces seizures and sponta-
subunits in discrete hippocampal regions of kindled animals. Brain Res
neous nonsynaptic bursting in vitro. Neurobiol Dis 2000; 7: 362-74.
67. Kral T, Erdmann E, Sochivko D, Clusmann H, Schramm J, Dietrich D.
43. Mathern GW, Bertram EH, Babb TL, Pretorius JK, Kuhlman PA,
Down-regulation of mGluR8 in pilocarpine epileptic rats. Synapse 2003;
Spradlin S, et al. In contrast to kindled seizures, the frequency of spon-
taneous epilepsy in the limbic status model correlates with greater
68. Jarvie PA, Logan TC, Geula C, Slevin JT. Entorhinal kindling perma-
aberrant fascia dentata excitatory and inhibitory axon sprouting, and
nently enhances Ca2+-dependent L-glutamate release in region inferior
increased staining for N-methyl-D-aspartate, AMPA and GABA re-
of rat hippocampus. Brain Res 1990; 508: 188-93.
ceptors. Neuroscience 1997; 77: 1003-19.
69. Sloviter RS. Permanently altered hippocampal structure, excitability,
44. Croucher MJ, Cotterell KL, Bradford HF. Amygdaloid kindling by re-
and inhibition after experimental status epilepticus in the rat: the ‘dor-
peated focal N-methyl-D-aspartate administration: comparison with
mant basket cell’ hypothesis and its possible relevance to temporal lobe
electrical kindling. Eur J Pharmacol 1995; 286: 265-71.
epilepsy. Hippocampus 1991; 1: 41-66.
45. McNamara JO, Russell RD, Rigsbee L, Bonhaus DW. Anticonvulsant
70. Sloviter RS. Possible functional consequences of synaptic reorganiza-
and antiepileptogenic actions of MK-801 in the kindling and electro-
tion in the dentate gyrus of entate-treated rats. Neurosci Lett 1992; 137:
shock models. Neuropharmacology 1988; 27: 563-8.
46. Stafstrom CE, Holmes GL, Thompson JL. MK801 pretreatment redu-
71. Shirasaka Y, Wasterlain CG. Chronic epileptogenicity following focal
ces kainic acid-induced spontaneous seizures in prepubescent rats. Epil-
status epilepticus. Brain Res 1994; 655: 33-44.
72. Gilbert ME. Potentiation of inhibition with perforant path kindling: an
47. Mazarati AM, Wasterlain CG. N-Methyl-D-asparate receptor antago-
NMDA-receptor dependent process. Brain Res 1991; 564: 109-16.
nists abolish the maintenance phase of self-sustaining status epilepti-
73. Milgram NW, Yearwood T, Khurgel M, Ivy GO, Racine RJ. Changes in
cus in rat. Neurosci Lett 1999; 265: 187-90.
inhibitory processes in the hippocampus following recurrent seizures
induced by systemic administration of kainic acid. Brain Res 1991; 551:
99. Menéndez de la Prida L, Sola RG, Pozo MA. Aspectos neurofisiológi-
cos in vivo e in vitro de la epilepsia. Rev Neurol 2002; 34: 430-7.
74. Kamphuis W, Lopes da Silva FH, Wadman WJ. Changes in local evok-
100. Hwa GG, Avoli M, Oliver A, Villemure JG. Bicucuclline-induced epi-
ed potentials in the rat hippocampus (CA1) during kindling epileptoge-
leptogenesis in the human neocrtex maintained in vitro. Exp Brain Res
75. Kapur J, Lothman EW. Loss of inhibition precedes delayed sponta-
101. McCormick DA. GABA as an inhibitory neurotransmitter in human
neous seizures in the hippocampus after tetanic electrical stimulation. J
cerebral cortex. J Neurophysiol 1989; 54: 782-806.
Neurophysiol 1989; 61: 427-34.
102. Gutnick, MJ, Connors BW, Prince DA. Mechanisms of neocortical epi-
76. Wu K, Leung LS. Increased dendritic excitability in hippocampal
leptogenesis in vitro. J Neurophysiol 1982; 48: 1321-35.
CA1 in vivo in the kainic acid model of temporal lobe epilepsy: a
103. Hwa GG, Avoli M. Excitatory synaptic transmission mediated by
study using current source density analysis. Neuroscience 2003; 116:
NMDA and non-NMDA receptors in the superficial/middle layers of
the epileptogenic human neocortex maintained in vitro. Neurosci Lett
77. Cossart R, Dinocourt C, Hirsch JC, Merchán-Pérez A, De Felipe J,
Ben-Ari Y, et al. Dendritic but not somatic GABAergic inhibition is
104. Staley KJ, Soldo BL, Proctor WR. Ionic mechanisms of neuronal exci-
decreased in experimental epilepsy. Nat Neurosci 2001; 4: 52-62.
tation by inhibitory GABA receptors. Science 1995; 269: 977-81.
78. Avoli M, Louvel J, Pumain R, Köhling F. Cellular and molecular mecha-
105. Kaila K, Lamsa K, Smirnov S, Taira T, Voipio J. Long-lasting GABA-
nisms of epilepsy in the human brain. Prog Neurobiol 2005; 77: 166-200.
mediated depolarization evoked by high-frequency stimulation in
79. Halliwell JV. M-current in human neocortical neurones. Neurosci Lett
pyramidal neurons of rat hippocampal slice is attributable to a net-
work-driven, bicarbonate-dependent K+ transient. J Neurosci 1997; 17:
80. McCormick DA, Williamson A. Convergence and divergence of neuro-
transmitter action in human cerebral cortex. Proc Natl Acad Sci U S A
106. Smirnov S, Paalasmaa P, Uusisaari M, Voipio J, Kaila K. Pharmacolo-
gical isolation of the synaptic and nonsynaptic components of the
81. Foehring RC, Waters RS. Contributions of low-thresholds calcium cur-
GABA-mediated biphasic response in rat CA1 hippocampal pyramidal
rent and anomalous rectifier (Ih) to slow depolarizations underlying
cells. J Neurosci 1999; 19: 9252-60.
burst firing in human neocortical neurons in vitro. Neurosci Lett 1991;
107. Louvel J, Papatheodoropoulos C, Siniscalchi A, Kurciewicz I, Pumain
R, Devaux, B, et al. GABA-mediated synchronization in the human
82. Lorenzon NM, Foehing RC. Relationship between repetitive firing and
cortex: elevations in extracellular potassium and presynaptic mecha-
afterhyperpolarizations in human neocortical neurons. J Neurophysiol
nisms. Neuroscience 2001; 105: 803-13.
108. Capogna M, Gahwiler BH, Thompson SM. Mechanism of m-opioid
83. Sayer RJ, Brown AM, Schwindt PC, Crill WE. Calcium currents in
receptor-mediated presynaptic inhibition in the rat hippocampus in vi-
acutely isolated human neocortical neurons. J Neurophysiol 1993; 69:
tro. J Physiol (Lond) 1993; 470: 539-58.
109. Babb TL, Brown WJ. Pathological findings in epilepsy. In Engel J Jr,
84. Cummins TR, Xia Y, Haddad GG. Functional properties of rat and hu-
ed. Surgical treatment of the epilepsies. New York: Raven Press; 1987.
man neocortical voltage-sensitive sodium currents. J Neurophysiol 1994;
110. Marco P, Sola RG, Pulido P, Alijarde MT, Sánchez A, Ramón y Cajal S,
85. Vreugdenhil M, Van Veelen CW, Van Rijes PC, Lopes da Silva FH,
et al. Inhibitory neurons in the human epileptogenic temporal neocor-
Wadman WJ. Effect of valproic acid on sodium currents in cortical
tex. An immunocytochemical study. Brain 1996; 119 (Pt 4): 1327-47.
neurons from patients with pharmaco-resistant temporal lobe epilepsy.
111. Ferrer I, Oliver B, Russi A, Casas R, Rivera R. Parvalbumin and cal-
bindin-D28k immunocytochemistry in human neocortical epileptic foci.
86. Avoli M, Olivier A. Electrophysiological properties and synaptic res-
ponses in the deep layers of the human epileptogenic neocortex main-
112. Ying Z, Babb TL, Comair YG, Bingaman W, Bushey M, Touhalisky K.
tained in vitro. J Neurophysiol 1989; 61: 589-606.
Induced expression of NMDAR2 proteins and differential expression
87. Avoli M, Mattia D, Siniscaldhi A, Perrealult P, Tomaiuolo F. Pharma-
of NMDAR1 splice variants in dysplastic neurons of human epileptic
cology and electrophysiology of a synchronous GABA-mediated po-
neocortex. J Neuropathol Exp Neurol 1998; 57: 47-62.
tential in the human neocortex. Neuroscience 1994; 62: 655-66.
113. González-Albó MC, Gómez-Utrero E, Sánchez A, Sola RG, DeFelipe
88. Menéndez de la Prida L, Benavides-Piccione R, Sola RG, Pozo MA.
J. Changes in the colocalization of glutamate ionotropic receptor sub-
Electrophysiological properties of interneurons from intraoperative
units in the human epileptic temporal lobe cortex. Exp Brain Res 2001;
spiking areas of epileptic human temporal neocortex. Neuroreport 2002;
114. Kortenbruck G, Berger E, Speckmann EJ, Musshoff U. RNA editing at
89. Cohen I, Navarro V, Clemenceau S, Baulac M, Miles R. On the origin
the Q/R site for the glutamate receptor subunits GLUR2, GLUR5, and
of interictal activity in human temporal lobe epilepsy in vitro. Science
GLUR6 in hippocampus and temporal cortex from epileptic patients.
90. Hwa, GG, Avoli M. The involvement of excitatory amino acids in neo-
115. Mirnics K, Middleton FA, Lewis DA, Levitt P. Analysis of complex
cortical epileptogenesis: NMDA and non-NMDA receptors. Exp Brain
brain disorders with gene expression microarrays: schizophrenia as a
disease of the synapse. Trends Neurosci 2001; 24: 479-86.
91. Avoli M, Hwa GC, Louvel J, Kurvewicz I, Pumain R, Lacaille JC.
116. Arion D, Sabatini M, Unger T, Pastor J, Alonso-Nanclares L, Balles-
Functional and pharmacological properties of GABA-mediated inhibi-
teros-Yáñez I, et al. Correlation of transcriptome profile with electrical
tion in the human neocortex. Can J Physiol Pharm 1997; 75: 526-34.
activity in the neocortex of temporal lobe epilepsy. Neurobiol Dis 2006
92. Deisz R. GABA(B) receptor-mediated effects in human and rats neo-
cortical neurones in vitro. Neuropharmacology 1999; 38: 1755-66.
117. Du F, Whetsell WO, Abou-Khalil B, Blumenkopf B, Lothman EW,
93. Strowbridge BW, Masukawa LM, Spencer DD, Sheperd GM. Hyperx-
Schwarcz R. Preferential neuronal loss in layer III of the entorhinal cortex
citability associated with localizable lesions in epileptic patients. Brain
in patients with temporal lobe epilepsy. Epilepsy Res 1993; 16: 223-33.
118. Lanerolle NC, Brines ML, Kim JH, Williamson A, Philips MF,
94. Williamson A, Patrylo PR, Lee S, Spencer DD. Physiology of human
Spencer DD. Neurochemical remodeling of the hippocampus in human
cortical neurons adjacent to cavernous malformations. Epilepsia 2003;
temporal lobe epilepsy. Epilepsy Res Suppl 1992; 9: 205-20.
119. Arellano JI, Muñoz A, Ballesteros-Yáñez I, Sola RG, DeFelipe J. His-
95. Schwartzkroin PA, Knowles WD. Intracellular study of human epilep-
topathology and reorganization of chandelier cells in the human epi-
tic cortex: in vitro maintenance of epileptiform activity? Science 1984;
leptic sclerotic hippocampus. Brain 2004; 127: 45-64.
120. Sutula T, Cascino G, Cavazos J, Parada I, Ramírez L. Mossy fiber sy-
96. Schwartzkroin PA, Haglund MM. Spontaneous rhythmic synchronous
naptic reorganization in the epileptic human temporal lobe. Ann Neu-
activity in epileptic hman and normal monkey temporal lobe. Epilepsia
121. Houser CR, Miyashiro JE, Swartz BE, Walsh GO, Rich JR, Delgado-
97. Köhling R, Lücke A, Straub H, Speckmann EJ, Tuxhorn I, Wolf P, et al.
Escueta AV. Altered patterns of dynorphin immunoreactivity suggest
Spontaneous sharp waves in human neocortical slices excised from
mossy fiber reorganization in human hippocampal epilepsy. J Neurosci
epileptic patients. Brain 1998; 121: 1073-87.
98. Köhling R, Höhling JM, Straub H, Kuhlmann D, Kuhnt U, Tuxhorn I,
122. Isokawa M, Levesque MF, Babb TL, Engel J Jr. Single mossy fiber
et al. Optical monitoring of neuronal activityduring spontaneous sharp
axonal systems of human dentate granule cells studied in hippocampal
waves in chronically epileptic human neocortical tissue. J Neurophysiol
slices from patients with temporal lobe epilepsy. J Neurosci 1993; 13:
123. Isokawa M. Remodeling dendritic spines of dentate granule cells in
epilepsy-associated brain tumors and in the perilesional epileptic cor-
temporal lobe epilepsy patients and the rat pilocarpine model. Epilep-
tex. Acta Neuropathol (Berl) 2001; 101: 449-59.
145. Fonseca CG, Green CR, Nicholson LF. Upregulation in astrocytic con-
124. De Felipe J. Chandelier cells and epilepsy. Brain 1999; 122: 1807-22.
nexin 43 gap junction levels may exacerbate generalized seizures in
125. Isokawa M, Avanzini G, Finch DM, Babb TL, Levesque MF. Physio-
mesial temporal lobe epilepsy. Brain Res 2002; 929: 105-16.
logic properties of human dentate granule cells in slices prepared from
146. Pacia SV, Ebersole JS. Intracranial EEG in temporal lobe epilepsy. J
epileptic patients. Epilepsy Res 1991; 9: 242-50.
Clin Neurophysiol 1999; 16: 399-407.
126. Vreugdenhil M, Hoogland G, Van Veelen CW, Wadman WJ. Persistent
147. Bragin A, Wilson CL, Straba RJ, Reddick M, Fried I, Engel J Jr. Inter-
sodium current in subicular neurons isolated from patients with tempo-
ictal high-frequency oscillations (80-500 Hz) in the human epileptic
ral lobe epilepsy. Eur J Neurosci 2004; 19: 2769-78.
brain: entorhinal cortex. Ann Neurol 2002; 52: 407-15.
127. Wozny C, Kivi A, Lehmann TN, Dehnicke C, Heinemann U, Behr J.
148. Wieser HG, Elger CE, Stodieck SRG. The foramen ovale electrode: a
Comment on ‘On the origin of interictal activity in human temporal
new recording method for the preoperative evaluation of patients suf-
lobe epilepsy in vitro’. Science 2003; 301: 463.
fering from mesio-basal temporal lobe epilepsy. Electroencephalogr
128. Bender RA, Soleymani SV, Brewster AL, Nguyen ST, Beck H, Mathern
GW, et al. Enhanced expression of a specific hyperpolarization-activat-
149. Sola RG, Hernando V, Pastor J, Navarrete EG, DeFelipe J, Alijarde
ed cyclic nucleotide-gated cation channel (HCN) in surviving dentate
MT, et al. Epilepsia farmacorresistente del lóbulo temporal. Explo-
gyrus granule cells of human and experimental epileptic hippocampus.
ración con electrodos del foramen oval y resultados quirúrgicos. Rev
129. Isokawa M, Levesque M, Fried I, Engel J Jr. Glutamate currents in
150. Pastor J, Hernando V, Domínguez-Gadea L, De Llano I, Meilán ML,
morphologically identified human dentate granule cells in temporal
Martínez-Chacón JL, et al. Impacto de la experiencia sobre los resulta-
lobe epilepsy. J Neurophysiol 1997; 77: 3355-69.
dos quirúrgicos en la epilepsia del lóbulo temporal. Rev Neurol 2005;
130. Mathern GW, Pretorius JK, Mendoza D, Leite JP, Chimelli L, Born
DE, et al. Hippocampal N-methyl-D-aspartate receptor subunit mRNA
151. Pastor J, Hernando V, Menéndez de la Prida L, Sola RG. Voltage sour-
levels in temporal lobe epilepsy patients. Ann Neurol 1999; 46: 343-58.
ces in mesial temporal lobe epilepsy recorded with foramen ovale elec-
131. Mathern GW, Pretorius JK, Kornblum HI, Mendoza D, Lozada A,
trodes. Clin Neurophysiol 2006 [submitted].
Leite JP, et al. Altered hippocampal kainate-receptor mRNA levels in
152. Thomas LB, Schmidt RP, Ward AA Jr. Observations on single units in
temporal lobe epilepsy patients. Neurobiol Dis 1998; 5: 151-76.
chronic cortical epileptic foci and in normal or strychninized cortex.
132. Mathern GW, Pretorius JK, Kornblum HI, Mendoza D, Lozada A,
Electroencephalogr Clin Neurophysiol Suppl 1955; 7: 478-80.
Leite JP, et al. Human hippocampal AMPA and NMDA mRNA levels
153. Wyler AR. Neuronal activity during seizures in monkeys. Exp Neurol
in temporal lobe epilepsy patients. Brain 1997; 120 (Pt 11): 1937-59.
133. Dietrich D, Kral T, Clusmann H, Friedl M, Schramm J. Reduced func-
154. Engel J Jr, Wilson C, Bragin A. Advances in understanding the process
tion of L-AP4-sensitive metabotropic glutamate receptors in human
of epileptogenesis based on patient material: what can the patient tell
epileptic sclerotic hippocampus. Eur J Neurosci 1999; 11: 1109-13.
us? Epilepsia 2003; 44 (Suppl 12): 60-71.
134. Olsen RW, Bureau M, Houser CR, Delgado-Escueta AV, Richards JG,
155. Staba RJ, Wilson CL, Bragin A, Fried I, Engel J Jr. Sleep states differ-
Möhler H. GABA/benzodiazepine receptors in human focal epilepsy.
entiate neuron activity recorded from human epileptic hippocampus,
Epilepsy Res 1992; 8 (Suppl): 383-91.
entorhinal cortex, and subiculum. J Neurosci 2002; 22: 5694-704.
135. Wolf HK, Spänle M, Müller MB, Elger CE, Schramm J, Wiestler OD.
156. Buzsaki G. Functions for interneuronal nets in the hippocampus. Can J
Hippocampal loss of the GABA receptor alpha 1 subunit in patients with
chronic pharmacoresistant epilepsies. Acta Neuropathol 1994; 88: 313-9.
157. Traub RD Whittington MA, Buhl EH. A possible role for gap junctions
136. Olsen RW, Avoli M. GABA and epileptogenesis. Epilepsia 1997; 38:
in generation of very fast EEG oscillations preceding the onset of, and
perhaps initiating, seizures. Epilepsia 2001; 42: 153-70.
137. Loup F, Wieser HG, Yonekawa Y, Aguzzi A, Fritschy JM. Selective al-
158. Wilson CL, Khan SU, Engel J Jr, Isokawa M, Babb TL, Behnke EJ.
terations in GABA receptor subtypes in human temporal lobe epilep-
Paired pulse suppression and facilitation in human epileptogenic hip-
pocampal formation. Epilepsy Res 1998; 31: 211-30.
138. Muñoz A, Méndez P, Álvarez-Leefmans FJ, DeFelipe J. 2004. Expres-
159. Beck H, Goussakov IV, Lie A, Helmstaedter C, Elger CE. Synaptic
sion of cation-chloride cotransporters NKCC and KCC2 in normal and
plasticity in the human dentate gyrus. J Neurosci 2000; 20: 7080-6.
epileptic hippocampus of humans. FENS 2004 [abstract 2, A197.2.385].
160. Rosenstein MT, Collins JC, De Luca CJ. A practical method for calcu-
139. Isokawa M. Decrement of GABA receptor-mediated inhibitory post-
lating the largest Lyapunov exponents from small data sets, Physica D
synaptic currents in dentate granule cells in epileptic hippocampus. J
161. Grassberger P, Procaccia I. Characterization of strange attractors. Phys
140. Vale C, Sanes DH. Afferent regulation of inhibitory synaptic transmis-
sion in the developing auditory midbrain. J Neurosci 2000; 20: 1912-21.
162. Schiff SJ, So P, Chang T, Burke RE, Sauer T. Detecting dynamical
141. Rivera C, Voipio J, Thomas-Crusells J, Li H, Emri Z, Sipila S, et al. Me-
interdependence and generalized synchrony through mutual prediction
chanism of activitydependent downregulation of the neuron-specific
in a neural ensemble. Phys Rev E 1996; 54: 6708-724.
K-Cl cotransporter KCC2. J Neurosci 2004; 24: 4683-91.
163. Lopes da Silva FH, Pijn JP, Wadman WJ. Dynamics of local neuronal
142. Furtinger S, Pirker S, Czech T, Baumgartner C, Ransmayr G, Sperk G.
networks: control parameters and state bifurcations in epileptogenesis.
Plasticity of Y1 and Y2 receptors and neuropeptide Y fibers in patients
with temporal lobe epilepsy. J Neurosci 2001; 21: 5804-12.
164. Pijn JP, Velis DN, Van der Heyden MJ, DeGoede J, Van Veelen CW,
143. Baraban SC, Tallent MK. Interneuron diversity series: interneuronal neu-
Lopes da Silva FH. Nonlinear dynamics of epileptic seizures on basis
ropeptides-endogenous regulators of neuronal excitability. TINS 2004;
of intracranial EEG recordings. Brain Topogr 1997; 9: 249-70.
165. Elger CE, Widman G, Andrzejak R, Dumpelmann M, Arnhold J, Grass-
144. Aronica E, Gorter JA, Jansen GH, Leenstra S, Yankaya B, Troost D.
berger P, et al. Value of nonlinear time series analysis of the EEG in
Expression of connexin 43 and connexin 32 gap-junction proteins in
neocortical epilepsies. Adv Neurol 2000; 84: 317-30. BASES FISIOPATOLÓGICAS DE LA EPILEPSIA DEL LÓBULOBASES FISIOPATOLÓGICAS DA EPILEPSIA DO LOBO TEMPORAL: ESTUDIOS EN HUMANOS Y ANIMALESTEMPORAL: ESTUDOS EM HUMANOS E EM ANIMAISResumen. Introducción. La epilepsia del lóbulo temporal (ELT) es Resumo. Introdução. A epilepsia do lobo temporal (ELT) é o tipo el tipo más frecuente de epilepsia farmacorresistente en humanos.mais frequente de epilepsia farmacorresistente em humanos. O tra-El tratamiento quirúrgico de estos pacientes permite estudios de grantamento cirúrgico destes doentes permite estudos de grande impor-importancia para conocer los mecanismos fisiopatológicos sub-tância para conhecer os mecanismos fisiopatológicos subjacentes.yacentes. Desarrollo. Se revisan algunos de los datos y teorías más
Desenvolvimento. Revêem-se alguns dos dados e teorias mais re-recientes sobre la fisiopatología de la ELT, tanto en modelos anima-centes sobre a fisiopatologia da ELT, tanto em modelos animais co-les como en humanos. Aunque existen similitudes electrofisiológi-mo em humanos. Embora existam semelhanças electrofisiológicas,cas, no hay una relación perfecta entre los datos hallados en mode-não existe uma relação perfeita entre os dados achados em modeloslos animales y en humanos, lo que dificulta la extrapolación de losanimais e em humanos, o que dificulta a extrapolação dos resulta-resultados. No obstante, ambos tipos de estudios sugieren una con-dos. No entanto, ambos os tipos de estudos sugerem uma considerá-siderable heterogeneidad en las alteraciones responsables de la epi-vel heterogeneidade nas alterações responsáveis da epilepsia, em-lepsia, si bien se acepta comúnmente que existe una remodelaciónbora seja comummente aceite que existe uma remodelação da exci-de la excitación glutamatérgica y la inhibición gabérgica que derivatação glutamatérgica e a inibição gabaérgica que resulta em hiper-en hiperexcitabilidad. El papel de la esclerosis mesial como procesoexcitabilidade. O papel da esclerose mesial como processo originá-originario de la ELT está cada vez más discutido. El desarrollo derio da ELT é cada vez mais discutido. O desenvolvimento de estudosestudios electrofisiológicos in vitro y la aplicación de técnicas conelectrofisiológicos in vitro e a aplicação de técnicas com maior po-mayor poder de resolución, como los registros con microelectrodosder de resolução, como os registos com microeléctrodos ou novaso nuevas herramientas matemáticas, pueden aportar importantesferramentas matemáticas, podem fornecer dados importantes aodatos al conocimiento fisiopatológico de este síndrome. Conclusio-
conhecimento fisiopatológico desta síndroma. Conclusões. O estu-
nes. El estudio multidisciplinar de la fisiopatología de la ELT en lado multidisciplinar da fisiopatologia da ELT na última década per-última década ha permitido aumentar el conocimiento sobre los pro-mitiu aumentar o conhecimento sobre os processos que subjazem àcesos que subyacen a la génesis de las crisis, su clínica y evolución.génese das crises, à sua clínica e evolução. Este conhecimento é deEste conocimiento es de gran importancia porque abre nuevas op-grande importância porque abre novas opções terapêuticas para aciones terapéuticas de la ELT. [REV NEUROL 2006; 42: 663-73]ELT. [REV NEUROL 2006; 42: 663-73]Palabras clave. EEG. Epilepsia del lóbulo temporal. Esclerosis me- Palavras chave. EEG. Epilepsia do lobo temporal. Esclerose me- sial. Kindling. Microelectrodos. Receptores de membrana. Regis-sial. Kindling. Microeléctrodos. Receptores da membrana. Regis-
INVESTIR EN PRIVATE EQUITY Assemblée Générale Ordinaire et Extraordinaire 30 juin 2011 Information sur les candidats aux fonctions de membre du Conseil de Surveillance Le mandat de onze des douze membres du Conseil de Surveillance vient à échéance à l’issue de la présente assemblée générale ordinaire et extraordinaire devant statuer sur les comptes clos le 31 déce
Correspondence Since population is a dynamic terised by diversity.4 This suggests that continuum of generations, there are the dis tinc tion between the “third age” issues in every age group to deal with and the “fourth age” (fi rst developed by information is based on forecasts. to secure healthy longevity as a whole. the histo r ical sociologist, Peter Laslett5) We declare that w
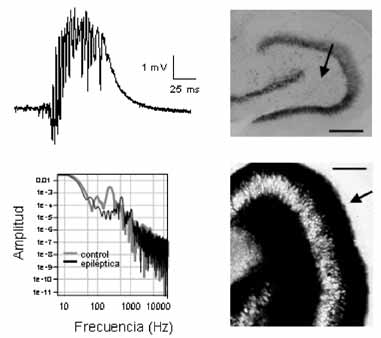 La alteración más típica del hipocampo
epiléptico es la esclerosis, caracterizada porla reinervación de los gránulos del giro denta-do por las fibras musgosas y la muerte neuro-nal en el hilus, zonas de CA3 y CA1 y las ca-pas superficiales de la corteza entorrinal (Figs.
La alteración más típica del hipocampo
epiléptico es la esclerosis, caracterizada porla reinervación de los gránulos del giro denta-do por las fibras musgosas y la muerte neuro-nal en el hilus, zonas de CA3 y CA1 y las ca-pas superficiales de la corteza entorrinal (Figs.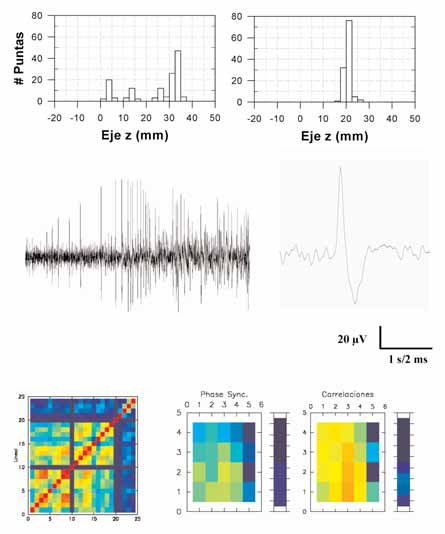 tes. El efecto despolarizante parece de-berse a un aumento anómalo de la con-
centración intracelular de Cl–, presumi-blemente debido a alteraciones de los co-
transportadores KCC2 y NKCC1, querevertiría la dirección de la corrienteinducida por la apertura de los canales
sión disminuida del cotransportador KCC2es una consecuencia de la desaferentiza-
ción de CA1 observada en la ELT [140,141], aunque también hay resultados si-milares en tejido no esclerótico [127].
ciones en la región mesial, cuya interpre-tación es aún incierta y se encuentran eninvestigación. Es el caso del incrementoen la expresión de los receptores Y2 y ladisminución de los receptores Y1 –am-bos del neuropéptido Y– en el hipocam-po de pacientes con ELT [142]. Como elneuropéptido Y regula la actividad de in-terneuronas y el flujo sináptico de Ca2+[143], las alteraciones descritas previa-mente podrían tener repercusión sobre laexcitabilidad del hipocampo. Otro inte-
resante mecanismo que se está estudian-do activamente es la participación de lasuniones eléctricas en el control de la ex-citabilidad. Se ha observado una eleva-ción de ARNm de conexina 43 en tejidoepiléptico [144,145], lo que indicaría queun incremento de la presencia de estaproteína podría modificar las propieda-des de sincronización del tejido.
tes. El efecto despolarizante parece de-berse a un aumento anómalo de la con-
centración intracelular de Cl–, presumi-blemente debido a alteraciones de los co-
transportadores KCC2 y NKCC1, querevertiría la dirección de la corrienteinducida por la apertura de los canales
sión disminuida del cotransportador KCC2es una consecuencia de la desaferentiza-
ción de CA1 observada en la ELT [140,141], aunque también hay resultados si-milares en tejido no esclerótico [127].
ciones en la región mesial, cuya interpre-tación es aún incierta y se encuentran eninvestigación. Es el caso del incrementoen la expresión de los receptores Y2 y ladisminución de los receptores Y1 –am-bos del neuropéptido Y– en el hipocam-po de pacientes con ELT [142]. Como elneuropéptido Y regula la actividad de in-terneuronas y el flujo sináptico de Ca2+[143], las alteraciones descritas previa-mente podrían tener repercusión sobre laexcitabilidad del hipocampo. Otro inte-
resante mecanismo que se está estudian-do activamente es la participación de lasuniones eléctricas en el control de la ex-citabilidad. Se ha observado una eleva-ción de ARNm de conexina 43 en tejidoepiléptico [144,145], lo que indicaría queun incremento de la presencia de estaproteína podría modificar las propieda-des de sincronización del tejido.