Tadalafil gehört zur Gruppe der PDE5-Hemmer und wirkt über eine hochselektive Blockade des Enzyms Phosphodiesterase Typ 5. Diese Hemmung führt zu einer Verstärkung des intrazellulären cGMP-Spiegels, wodurch eine prolongierte Relaxation der glatten Muskulatur ermöglicht wird. Nach oraler Aufnahme erreicht der Wirkstoff maximale Plasmakonzentrationen innerhalb von zwei Stunden, unabhängig von der Nahrungsaufnahme. Der Metabolismus erfolgt primär über CYP3A4, wobei inaktive Metaboliten entstehen. Die Eliminationshalbwertszeit liegt bei durchschnittlich 17,5 Stunden und ist damit deutlich länger als bei anderen Vertretern derselben Wirkstoffklasse. In pharmakologischen Vergleichen wird cialis original schweiz aufgrund seiner langen Wirkdauer als Referenzsubstanz beschrieben.
Tri conference template
Genetic diversity in Australian canola and implications for future breeding
Wallace A. Cowling
School of Plant Biology, The University of Western Australia, 35 Stirling Highway, Crawley, WA 6009, AUSTRALIA Email:
Abstract Australian canola breeders have made excellent progress in developing blackleg resistant and high quality canola varieties in approximately 5 breeding cycles from 1970 – 2000, averaging 6 years/cycle in a closed population. Of the original 18 parents in 1970, which included Brassica napus from Europe and Asia and two B. juncea varieties, 11 ancestor varieties now contribute 98.7% of the pedigree composition of the core population. The inbreeding coefficient at the end of the fifth cycle was 0.21, which represents 21% cumulative loss of alleles through random genetic drift. The effective population size of the core population was relatively low (~11), but “immigrant” varieties have increased this in recent years. Two examples will be used to demonstrate the importance of new genetic diversity for future genetic progress: adaptation to low rainfall environments, and improved polygenic blackleg resistance. Evidence exists for a slow decay in polygenic blackleg resistance which will need to be countered by breeding with new genetic diversity for improved resistance. Selection of early flowering plants is complicated by the partially out-crossing nature of B. napus, and the use of open-pollinated (OP) pedigree breeding systems in Australia over the past 30 years. Outcrossing in an OP breeding system can delay or prevent the selection of early-flowering lines that are free of later-flowering contaminants. However, protection of plants against outcrossing in a pedigree breeding system is very costly. New technologies such as doubled haploidy (DH) may improve outcomes when breeding with new sources of genetic diversity. Heritability among DH lines is high, and the net cost of pollination control is less than a pedigree breeding system. Major challenges exist for the Australian canola industry: to access global genetic resources and increase effective population size while improving local adaptation; and to adequately reward investment in canola breeding for the wheatbelt region. Keywords Oilseed rape, drought tolerance, recurrent selection, inbreeding, canola pedigrees Introduction Canola, or double-low oilseed rape (Brassica napus L.), is an important oilseed crop in southern Australia where it is grown mostly in the higher rainfall regions with relatively secure rainfall (>450 mm average annual rainfall). Breeding for low rainfall environments has been less successful, and canola is almost absent from areas averaging less than 325 mm average annual rainfall (Oilseeds WA, 2005). Early B. napus ‘rapeseed’ varieties introduced from Europe or Canada prior to 1970 were adapted to long season, high rainfall regions in Australia, but had extreme blackleg susceptibility – a combination that was fatal for the oilseed rape industry, which almost disappeared until the 1990s. Australian canola breeders have successfully improved yield, adaptation, blackleg resistance and seed quality (low erucic acid and low glucosinolates, known as “double low” or canola quality) since 1970 (Salisbury and Wratten 1999), such that the production of canola exceeded 1.6 million tonnes by 1998/99 (Colton and Potter 1999). Australian canola breeding has undergone several cycles of recurrent selection in a closed population since 1970, with new “immigrant” varieties appearing in pedigrees of the most recent cultivars. B. napus is predominantly self-pollinating but has an open flower that promotes cross-pollination, with up to 30% outcrossing among plants in close proximity in some situations (Rakow and Woods 1987). The breeding system used in Australia since 1970 has been an open-pollinated (OP) pedigree selection method. Many breeders in Europe and Canada use pollination bags to protect plants from outcrossing during pedigree selection. Bagging is a very expensive exercise but has certain benefits, especially for the selection of early maturing lines that are free of contamination from later maturing types. The cost of
2006 "Ground breaking stuff"
Proceedings of the 13th ASA Conference, 10-14 September 2006, Perth, Western Australia. Web site www.agronomy.org.au
bagging in a pedigree breeding system is extremely high, and alternative breeding methods such as doubled haploidy (DH) or single seed descent may be preferred for selection of early maturity. This will be discussed in this paper in the context of selecting canola for adaptation to low rainfall environments. Canola breeders in Australia have been very successful in developing blackleg resistant varieties adapted to medium-high rainfall regions. This paper evaluates genetic progress in this closed population over 30 years, including the efficiency of the breeding method, coefficients of inbreeding and ancestry, the potential decay in polygenic blackleg resistance over time, and the potential for broadening the adaptation of canola in current or future germplasm. The paper addresses the question of why canola is not grown widely in low rainfall cropping zones, and why most growers and agronomists consider canola to be a drought sensitive crop, compared with other species such as B. juncea. There are very good prospects for low rainfall canola breeding, but sufficient genetic diversity must be present in the breeding material and adequate incentive to breed this low-yielding but important Australian agricultural environment. Methods Pedigrees of Australian canola cultivars released between 1970 and 2004 were obtained from Salisbury and Wratten (1999) and the). These were then plotted to demonstrate pedigree connections, based on year of release of each cultivar, to the ancestor varieties used in 1970 (Fig. 1). Pedigree trees were used to calculate the percent contribution of each ancestor variety in each released cultivar, and the frequency of occurrence in pedigrees of cultivars released from 1995 to 2002 (Table 1). The number of crossing cycles from 1970 to the year of release of each variety was calculated from the average of the male and female side of each pedigree (Table 1). Inbreeding coefficient (F) and coefficient of ancestry or parentage (f) were calculated for representative cultivars from 1995 to 2002 following the methods outlined in Falconer and Mackay (1996), with the following modifications: the parents in each cross were assumed to be homozygous, and therefore no adjustment was made for F of common ancestors, but the contribution to F or f for each relationship path was increased by a factor of ½. This accounted for the inbred and homozygous condition of the common ancestor, which in animals or outcrossing plants would be heterozygous. The overall degree of “inbreeding” in the population over t cycles of recurrent selection was based on the formula for inbreeding coefficient over generations in a closed population (Falconer and Mackay, 1996):
where ΔF is the rate of inbreeding, which in a random mating population is related to effective population size (N ) as follows (Falconer and Mackay, 1996):
F is equivalent to the cumulative effect of random drift (Falconer and Mackay 1996).
Blackleg resistance of Australian canola cultivars is assessed by private and public breeders in a national testing scheme that is co-ordinated by the National Brassica Improvement Program (NBIP). Collaborators assess percentage stand decline due to blackleg disease on current and older cultivars with standard control varieties nominated by the NBIP, in replicated trials at 5-10 locations across southern Australia each year. Predicted values of percentage stand decline for genotypes across years and sites are converted into a scale from 0 (very susceptible) to 9 (very resistant), and these NBIP cultivar blackleg scores are published by the Canola Association of Australia ( Published and unpublished cultivar blackleg scores from the NBIP were used to assess “mature” blackleg scores of each canola variety, where “mature” indicates the NBIP score for a cultivar after a minimum of 5 years in the NBIP system. The average change in blackleg score per year was assessed by subtracting the mature blackleg score from the score in the year after release (excluding the preliminary score in the year of release), and dividing by the number of years. The average change in blackleg score per year was recorded for a variety only if there was a minimum of four years’ data after the preliminary assessment in the year of release.
2006 "Ground breaking stuff"
Proceedings of the 13th ASA Conference, 10-14 September 2006, Perth, Western Australia. Web site www.agronomy.org.au
2006 "Ground breaking stuff"
Proceedings of the 13th ASA Conference, 10-14 September 2006, Perth, Western Australia. Web site www.agronomy.org.au
2006 "Ground breaking stuff"
Proceedings of the 13th ASA Conference, 10-14 September 2006, Perth, Western Australia. Web site www.agronomy.org.au
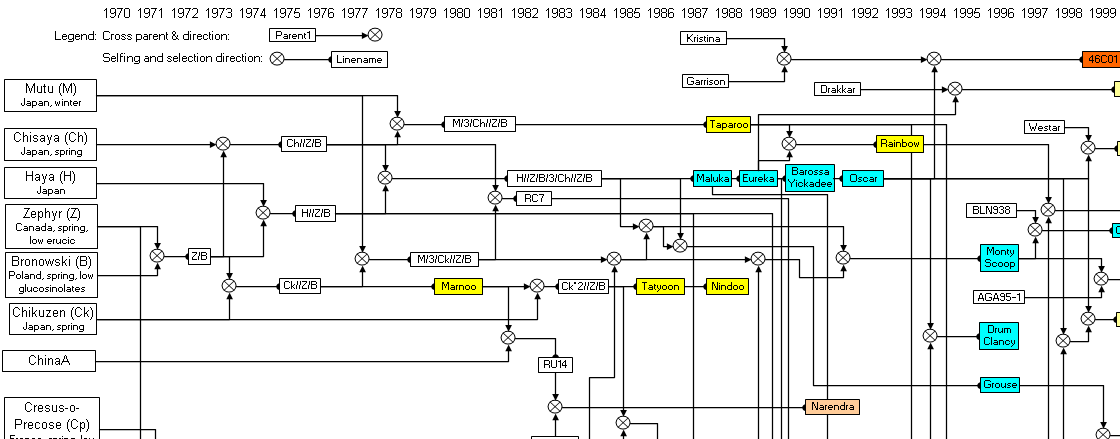
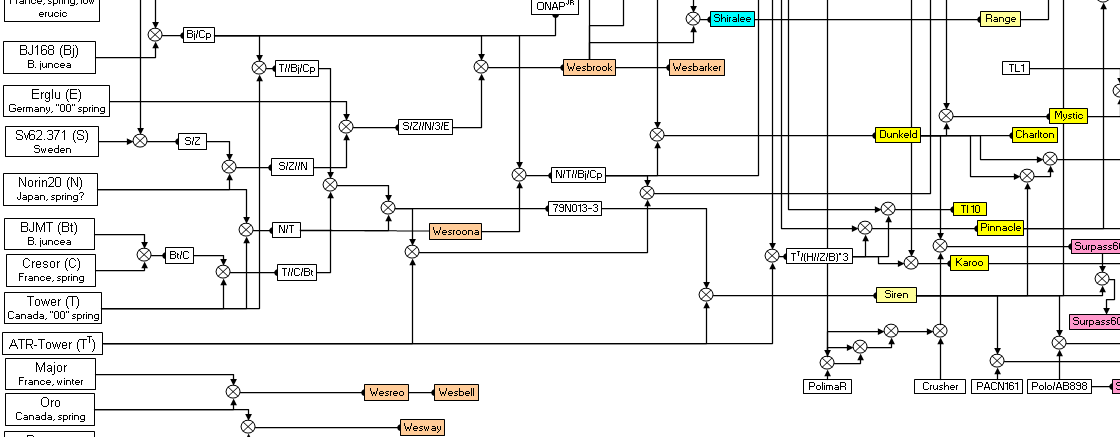
Figure 1. Pedigrees of selected Australian canola varieties 1970 to 2004 (from Wratten and Salisbury, 1999). Varieties are not listed if their pedigrees are uncertain (e.g., Rivette, Ripper, Purler, Lantern, AV-Sapphire, AG-Emblem, AG-Comet), hybrid or Clearfield (imidazolinone-tolerant).
Table 1. Pedigree analysis and blackleg resistance of canola varieties released 1995-2002, from the closed population derived from 18 ancestor varieties (used in original crossing cycle c. 1970), for which parent information is published (Salisbury and Wratten 1999, and IP Australia PBR Office). Varieties that are sister lines are combined (e.g. Drum-Clancy) and selections with the same pedigree are included only in the first instance (e.g. Pinnacle is included, ATR-Beacon is excluded). Backcrosses to the same parent are counted as one crossing cycle.
2006 "Ground breaking stuff"
Proceedings of the 13th ASA Conference, 10-14 September 2006, Perth, Western Australia. Web site www.agronomy.org.au
Grace Hyden Castle %contribution occurrence in
1Mature blackleg scores are the most recent blackleg resistance scores in published or unpublished data from Canola Association of Australia (http://www.canolaaustralia.com) or the National Brassica Improvement Program. Rating scale 0-9 where 0 = very susceptible and 9 = very resistant. Mature blackleg rating is assessed after a minimum of 5 years from release. 2Change in blackleg score per year in blackleg resistance units, from 2000 to most recent rating, not including the score in year of release (minimum 4 years data).
2006 "Ground breaking stuff"
Proceedings of the 13th ASA Conference, 10-14 September 2006, Perth, Western Australia. Web site www.agronomy.org.au
Results Effective population size and inbreeding Of the 18 parents used to begin Australian canola breeding in 1970 or thereabouts, Major, Oro and Ramses were used in the release of just three varieties, Wesreo (1978), Wesway (1979) and Wesbell (1980), and these varieties did not appear in pedigrees of subsequent varieties (Fig. 1). Another parent, ChinaA, was used only in the development of Narendra (1991), and this variety did not appear in pedigrees of subsequent varieties. Two varieties, Tower and ATR-Tower (triazine herbicide tolerant), were considered identical from a pedigree relationship point of view. Most varieties in the late 1990s were based on 14 ancestral varieties. The contribution of the ancestor parents in Australian canola varieties released between 1995 and 2002 are listed in Table 1. Two of these parents (BJMT and Cresor) appeared in only three of the varieties, and at less than 1% in each variety. Another variety, Sv62.371, contributed less than 1% on average across the 14 varieties. Approximately 98.7% of Australian canola germplasm in the late 1990s was derived from 11 ancestor parents, and four of these, Chisaya, Haya, Zephyr and Bronowski, contributed more than 50% of the ancestry on average across all varieties. The effective population size (N ), based on the
number of ancestors that survived in pedigrees of varieties in Table 1 at ≥1% of the ancestry, was estimated to be no greater than 11 from 1970 - 2000. While this value can, in theory, rise in this closed population following the initial “bottleneck” of introductions in 1970, this can only be achieved if large numbers of cross progeny contribute to crossing in future generations and there is no evidence from released varieties that this has occurred. The average number of recurrent selection cycles since 1970 in varieties released from 1995 – 1998 was 4.3, and in varieties released from 2000 – 2002 was 5.1 (Table 1). From this it was concluded that the closed population has undergone approximately 5 cycles during 30 years, at an average cycle time of 6 years from cross to cross. Based on N = 11, the of the population was predicted to be 0.21 after 5 cycles of recurrent selection,
which represents 21% cumulative loss of alleles through random genetic drift (Falconer and Mackay 1996). Individual varieties had inbreeding coefficients between 0.018 (Mystic) and 0.344 (ATR-Grace) (Table 1). The coefficient of ancestry of Monty (1996) Mystic (1998) was estimated to be 0.127, for Monty (1996) ATR-Grace (2001) was 0.299, and for ATR-Grace (2001) AG-Castle (2002) was 0.371. These values are increasing over time, and this indicates that inbreeding is increasing over time, as parents in each cycle are becoming more closely related. For comparison, full-sib animal mating results in a coefficient of ancestry of 0.25, and the mean coefficient of parentage of pairs of cultivars in two-row barley cultivars in North America averaged 0.19, and two- and six-row cultivars were on average unrelated (Martin et al. 1991). What is the relevance of coefficients of inbreeding (F) and ancestry (f) in a self-pollinating breeding system? The coefficient of ancestry can be used to select parents for crossing – if fine tuning of a trait is needed, closely related parents may be chosen. If new combinations of alleles are sought, then distantly related parents should be selected for crossing. “Inbreeding” takes on a new meaning in self-pollinating species – inbreeding depression is not likely to occur, as most lethal or detrimental mutations have been removed. The relatively small N of the Australian canola breeding pool means that genetic drift is
continually removing potentially valuable alleles. The value F = 0.21 may be interpreted as a 21%
random loss of alleles after five cycles of recurrent selection. While genetic progress is possible in the short term, new genetic material must be brought in to ensure future genetic progress. New “immigrants” began to appear in pedigrees of released varieties in Australia in the late 1990s from the breeding programs of Pacific Seeds Pty Ltd, AgSeed Research Pty Ltd, and Pioneer Hi-Bred Australia Pty Ltd (Fig. 1). This indicates that new genetic diversity is entering the Australian canola breeding gene pool. Improvement in blackleg resistance Significant genetic progress for blackleg resistance is apparent in the closed population as demonstrated by the moderate to high resistance of varieties released in the late 1990s and early 2000s (Table 1). The
2006 "Ground breaking stuff"
Proceedings of the 13th ASA Conference, 10-14 September 2006, Perth, Western Australia. Web site www.agronomy.org.au
blackleg resistance scores in Table 1 are the most recent values available for each variety, or the mature resistance score, and in many cases are 0.5 or 1.0 units lower than the values recorded in the year after release of the variety (see Methods and following paragraphs). Mature resistance scores are conservative and represent the field resistance of a variety after a minimum of 5 years from the date of commercial release. While the resistance ratings on ancestor varieties are not documented, it may be assumed that most of the European and Canadian ancestor lines were very susceptible (0-2 ratings) and some of the Japanese lines had intermediate ratings (possibly 3-5). The B. juncea ancestor lines may have had high resistance ratings, but none of the major resistance genes from B. juncea were transferred to B. napus varieties despite attempts to do so by interspecific crossing and development of major gene resistance B. napus line ONAPJR. The advanced generations of ONAPJR lines continued to segregate for major gene resistance and were not stable (Roy 1984). If the assumption is made that the average blackleg resistance score of ancestor varieties in 1970 was 2.0, and the average mature score in the advanced population in 2000 was 6.0 (Table 1), then the net resistance in the closed population rose by approximately 0.8 resistance units each cycle, or +0.13 resistance units per year. This is the net rate of improvement since 1970, taking into account any negative component due to the apparent fall in resistance of Australian cultivars in the years following release. The average change in blackleg resistance scores per year following release for cultivars in Table 1 ranged from 0.00 (Clancy) to -0.29 (Mystic). Data for 33 Australian canola varieties, which met the minimum requirements for inclusion, indicate that the blackleg resistance scores of canola changed by an average of -0.13 units per year after release, with a range from +0.10 to -0.29 (Table 2). This average is highly significantly different from zero (Table 2), and it is concluded that Australian canola varieties slowly lose polygenic resistance in the years following release. Nevertheless, it is clear that canola breeders have more than kept ahead of this erosion of polygenic resistance through genetic improvements in resistance in the closed breeding population. The net resistance improvement (the sum of genetic improvement and genetic erosion of resistance) over 30 years is +0.13 units per year. However, new genetic variation will need to be added to the closed population to maintain the current rate of genetic improvement in polygenic blackleg resistance into the future. There is strong circumstantial evidence that transgressive segregation has occurred for blackleg resistance over and above the levels of resistance found in the ancestor varieties. This improvement of polygenic blackleg resistance over 30 years by Australian canola breeders is a major success story, and demonstrates the power of recurrent selection in relatively small populations. However, this must be tempered by the knowledge that new systems of breeding may be needed to maintain the net improvement rate in blackleg resistance by expanding the genetic base. Adaptation to low rainfall environments Very little canola is grown in low rainfall regions (<325 mm) of Western Australia (Oilseeds WA 2005). Adaptation to this environment will require the development of new genotypes not yet seen in the core population of Australian canola breeding, with alleles that improve seed yield in short seasons with frequent periods of severe drought stress at the end of the season. Improved yield and adaptation will be the result of a complex combination of minor alleles and the best source of these alleles will be difficult to anticipate. It is possible that alleles for higher yield in low rainfall environments have been lost by genetic drift in the core population, as a result of breeding in relatively “safe” environments of south-eastern Australia over 30 years. If growing conditions are favourable and soil fertility not limiting, as occurs most often in primary sites of breeding programs, shorter and early maturing types will be discriminated against as a natural consequence of shading and poorer performance compared with later maturing larger plants. The selection of early lines in Australian OP breeding systems is complicated by several factors. Early maturing plants will tend to be out-competed and, without protection from outcrossing (Rakow and Woods 1987), they will inevitably have high levels of cross-pollination from surrounding mid- and late-maturing types. The seed derived from OP single plants will be segregating for flowering time, and
2006 "Ground breaking stuff"
Proceedings of the 13th ASA Conference, 10-14 September 2006, Perth, Western Australia. Web site www.agronomy.org.au
heterozygous, and plots in following years derived from these plants will have mixed maturity. Further reselection will be required, resulting in a long delay or failure in breeding of early varieties. However, the cost of bagging plants through the several generations of an OP pedigree breeding system is very expensive. New technologies may help to solve this dilemma. Doubled haploidy (DH) allows a random selection of genotypes from a cross to be preserved as primary homozygous plants, ready for selection. If the primary DH plant, and perhaps the next generation, is protected from outcrossing, there should be sufficient seed to select the small proportion of lines that are worthy of further consideration, and to put aside pure homozygous seed for future pure seed increase if it is warranted. This means that pollination protection should cost less than a pedigree breeding system, where selection of single plants may occur over several generations. Table 2. Change in blackleg resistance scores and mature blackleg score (bold) for 33 Australian Brassica napus cultivars.
2006 "Ground breaking stuff"
Proceedings of the 13th ASA Conference, 10-14 September 2006, Perth, Western Australia. Web site www.agronomy.org.au
2000 2001 2002 2003 2004 2005 2006 Change1 Years per year
Average change in blackleg score per year:
Probability average is different from zero:
1Change in blackleg score is calculated from the mature blackleg score (in bold) minus the first score after the preliminary score in year of release (preliminary score in italics). This is divided by the number of years separating the two scores to give the change in blackleg score per year. A minimum of four years' data were required to assess "change per year" and 33 varieties met this criterion.
DH lines express strong additive genetic variance and therefore heritability is high in early generations after production of the first DH plant – selections for early maturity will “breed true” for early maturity. In contrast, heritability is low in early generations of a pedigree breeding system as a result of heterozygosity, and early generation lines do not breed true, especially if they are exposed to continual cross-pollination. DH technology should allow selection of early maturing lines from a cross, more quickly, more confidently and more efficiently than with a pedigree breeding system. Very early maturing DH-derived cultivar Trilogy (Fig. 1) was selected and released for extremely low rainfall environments within 4 years of crossing. Conclusion Canola breeding in Australia from 1970 to 2000 was remarkable on the world stage for successful breeding of blackleg resistance by transgressive segregation from diverse ancestral parents, for development of high seed quality and for adaptation to Australian environments. However, there are now signs of loss of genetic diversity, and the coefficient of parentage between pairs of Australian canola
2006 "Ground breaking stuff"
Proceedings of the 13th ASA Conference, 10-14 September 2006, Perth, Western Australia. Web site www.agronomy.org.au
varieties in 2000 (>0.3) is higher than between pairs of North American two-row barley cultivars prior to 1990 (0.19) (Martin et al. 1991). New genetic diversity is required to promote future genetic progress in polygenic blackleg resistance and broader adaptation to the Australian wheatbelt. New methods of introgression of new alleles should be investigated; and new biotechnologies such as DH technology may aid the development of early maturing types with improved blackleg resistance and adaptation to low rainfall environments. These changes have already started. New “immigrant” varieties have appeared in pedigrees of recent varieties, and new low rainfall varieties are emerging (Fig. 1). The breeding principles of the RIPE system in barley should be considered for introgression of new alleles and expansion of the effective population size in Australian canola (Kannenberg and Falk 1995). More efforts are needed to expand the Australian canola gene pool and to take advantage of genetic improvements outside of Australia. The RIPE system deliberately restricts the rate of introgression in order to preserve local adaptation. This allows a balance between the rate of introgression of new alleles and improvements in local adaptation. There are no canola breeding programs outside of Australia with germplasm that is adapted to Australian conditions. Any imported germplasm will be cross-bred with local types to introgress valuable alleles. With new genetic diversity and genetic improvements for low rainfall in the next few years, B. napus canola should no longer be considered a drought-sensitive crop by Australian growers and agronomists. There are economic issues for canola breeding that also need to be addressed by industry. Australia needs to develop a system that adequately rewards investment in canola breeding for the lower rainfall regions, and which also meets the needs of growers. Acknowledgements The author appreciates the help of D Falk in reviewing the manuscript and critically assessing the calculation of coefficients of inbreeding and ancestry, and the National Brassica Improvement Program for providing blackleg resistance data from 2000 – 2005. This paper is the result of academic research at The University of Western Australia; the author’s salary for academic research was funded by the Export Grains Centre Ltd. References Colton, B, and Potter, T (1999). History. In Canola in Australia: the First 30 Years. Eds PA Salisbury,
TD Potter, G McDonald, AG Green. pp. 1-4, Organising Committee of the 10th International Rapeseed Congress.
Falconer, DS, and Mackay, TFC (1996). Introduction to Quantitative Genetics. Pearson Education
Kannenberg, LW, and Falk, DE (1995). Models of activation of plant genetic resources for crop breeding
programs. Canadian Journal of Plant Science 75, 45-53
Martin, JM, Carter, Jr, TE, and Burton, JW. (1991). Diversity among North American spring barley
cultivars based on coefficients of parentage. Crop Science 31, 1131-1137.
Oilseeds WA (2005). Growing western canola in the low rainfall zone (<325 mm) update 2005.
Pamphlet produced by The Oilseeds Industry Association of WA Inc., for Crop Updates, Perth WA, February 2005.
Rakow, G, and Woods, DL (1987). Outcrossing in rape and mustard under Saskatchewan prairie
conditions. Canadian Journal of Plant Science 67, 147-151.
Roy, NN (1984). Interspecific transfer of Brassica juncea-type high blackleg resistance to Brassica
Salisbury, PA, and Wratten, N (1999). Brassica napus breeding. In Canola in Australia: the First 30
Years. Eds PA Salisbury, TD Potter, G McDonald, AG Green. pp. 29-35, Organising Committee of the 10th International Rapeseed Congress.
2006 "Ground breaking stuff"
Proceedings of the 13th ASA Conference, 10-14 September 2006, Perth, Western Australia. Web site www.agronomy.org.au
Nucleo™ ingredient list and nutritional information as it appears on the packaging. Both these lists are approved by the South African Directorate Food control. This product is registered as a complimentary medicine with the Medicines Control Counsel of South Africa. (Registration number 421750) Ingredients Pre cooked full yellow maize, soya milk powder, fructose, nature identical apple flavo
Micro Log™ Plus A wireless data logging system for remotemonitoring of up to 200 data loggers The MicroLogTM Plus is a unique system based on the MicroLog – a mini temperature and humidity data logger. The MicroLog Plus allows users to communicate with up to 200 MicroLogs and receive all real-time measurements on one computer. The MicroLog Plus in
 2006 "Ground breaking stuff"
Proceedings of the 13th ASA Conference, 10-14 September 2006, Perth, Western Australia. Web site www.agronomy.org.au
Figure 1. Pedigrees of selected Australian canola varieties 1970 to 2004 (from Wratten and Salisbury, 1999). Varieties are not listed if their pedigrees are uncertain (e.g.,
2006 "Ground breaking stuff"
Proceedings of the 13th ASA Conference, 10-14 September 2006, Perth, Western Australia. Web site www.agronomy.org.au
Figure 1. Pedigrees of selected Australian canola varieties 1970 to 2004 (from Wratten and Salisbury, 1999). Varieties are not listed if their pedigrees are uncertain (e.g.,