Tadalafil gehört zur Gruppe der PDE5-Hemmer und wirkt über eine hochselektive Blockade des Enzyms Phosphodiesterase Typ 5. Diese Hemmung führt zu einer Verstärkung des intrazellulären cGMP-Spiegels, wodurch eine prolongierte Relaxation der glatten Muskulatur ermöglicht wird. Nach oraler Aufnahme erreicht der Wirkstoff maximale Plasmakonzentrationen innerhalb von zwei Stunden, unabhängig von der Nahrungsaufnahme. Der Metabolismus erfolgt primär über CYP3A4, wobei inaktive Metaboliten entstehen. Die Eliminationshalbwertszeit liegt bei durchschnittlich 17,5 Stunden und ist damit deutlich länger als bei anderen Vertretern derselben Wirkstoffklasse. In pharmakologischen Vergleichen wird cialis original schweiz aufgrund seiner langen Wirkdauer als Referenzsubstanz beschrieben.
4360036 779.782
Christoph E. Nabholz á Dave Speijer á Andre Schneider
Chloramphenicol-sensitive mitochondrial translationin Trypanosoma brucei
Received: 30 January 1999 / Accepted: 17 February 1999
Abstract We developed an in organello system to label
rRNAs among all eukaryotes and may therefore serve as
newly synthesized mitochondrially encoded proteins of
a system for investigation of the minimal requirements
Trypanosoma brucei. Highly puri®ed mitochondria,
for a protein synthesis machinery (Benne and Sloof
prepared under isotonic conditions, were incubated with
1987). Third, unlike mitochondrial translation in most
radioactive methionine and cysteine in a suitable trans-
other species, mitochondrial protein synthesis in try-
lation buer. Analysis of mitochondrial extracts on
panosomatids relies exclusively on tRNAs that are im-
TRIS-Tricine gels revealed a subset of labeled, NP-40-
ported from the cytosol (Schneider 1994).
insoluble proteins. The labeling of these proteins was
The problem of mitochondrial translation in trypan-
resistant to the cytosol-speci®c translation inhibitor
osomatids has proved to be dicult to analyze. Attempts
cycloheximide. The proteins, however, were not labeled
to purify mitochondrial ribosomes have been only par-
in the presence of chloramphenicol or erythromycin,
tially successful (Shu and GoÈringer 1998). Cytochrome c
inhibitors of prokaryotic type translation, or puromycin,
oxidase from Crithidia fasciculata has been puri®ed.
a general translation inhibitor. These results indicate
However, all the putative mitochondrially encoded sub-
that isotonically isolated mitochondria of T. brucei are
units were refractory to protein sequencing (Speijer et al.
1996; Brekk et al. 1997). The best evidence for mito-chondrial translation was found in a number of studiesusing antibodies raised against synthetic peptides of pu-tative mitochondrially encoded proteins that detected
signals of the expected molecular weights in the mito-chondrial fraction (Shaw et al. 1989; Beattie and Howton
Mitochondrial translation in trypanosomatids is of great
1996). In addition, in organello labeling experiments in
interest for at least three reasons. First, many RNAs
C. fasciculata identi®ed a number of putative mito-
that are synthesized in trypanosomatid mitochondria
chondrially encoded proteins. However, no information
undergo RNA editing by uridylate insertions and dele-
was provided as to whether the observed translation was
tions to become functional mRNAs. In recent years,
sensitive to inhibition by prokaryotic type translation
much has been learned concerning the mechanism of
inhibitors (Tittawella 1998). In the present study we used
RNA editing (Sollner-Webb 1996). However, nothing is
an in organello labeling approach for direct demonstra-
known about one important aspect of the problem,
tion of mitochondrial translation in Trypanosoma brucei.
whether and, if so, how fully edited mRNAs are trans-lated within mitochondria. Second, mitochondrial ribo-somes of trypanosomatids contain the shortest known
Procyclic wild-type and transformed Trypanosoma brucei, stock
University of Fribourg, Institute of Zoology, PeÂrolles,
427, were grown in SDM-79 medium supplemented with 5% fetal
e-mail: [email protected], Tel.: +41-26-300-8894,Fax: +41-26-300-9741
Protein Research Facility Amsterdam,E. C. Slater Instituut BioCentrum Amsterdam,
Mitochondria were isolated using sterilized buers and equipment
as described elsewhere (Hauser et al. 1996), except that a low-speed
spin (300 g) was routinely performed before loading of the Ny-codenz gradients. The absence of intact T. brucei cells and bacterialcontamination in isolated mitochondrial preparations was con-®rmed by light microscopy. In organello translations were per-formed using 400 lg of isolated mitochondria each in 100 ll oftranslation buer [20 mM TRIS-HCl (pH 7.4), 30 mM KH2PO4,0.6 M sorbitol, 5 mM succinate, 50 mM KCl, 20 mM MgSO4,12 mM creatine phosphate, creatine phosphokinase at 0.16 mg/ml,4 mM ATP, 0.5 mM GTP, 5 mM NADH, fatty-acid-free bovineserum albumin at 2.5 mg/ml, and 1 mM of all biological aminoacids except methionine and cysteine] containing 1 lM (approxi-mately 150 lCi) of a mixture of 35S-labeled methionine and cys-teine (Pro-mix, Amersham, Switzerland). The reactions wereincubated for 90 min at 25 °C, and mitochondria were reisolatedby centrifugation for 5 min at 5,200 g at 4 °C. The resulting pelletswere extracted with 100 ll of 50 mM potassium phosphate (pH7.5) containing 0.5% NP-40. Subsequently, the samples were cen-trifuged at 15,000 g for 15 min, and the pellet was solubilized underconstant mixing for 2 h at 37 °C in 50 ll of 0.28 M TRIS-HCl (pH6.8), 2.8% sodium dodecyl sulfate, 5% (v/v) glycerol, and 5% (v/v)b-mercaptoethanol. All reactions were analyzed on TRIS-Tricinegels (SchaÈgger and von Jagow 1987), which were ®rst stained withCoomassie brilliant blue to check for equal loading and then pro-cessed for ¯uorography.
In organello translation reactions were incubated in the presence
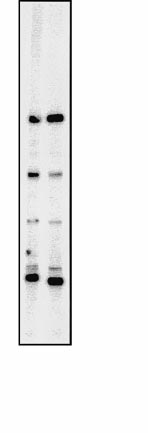
Fig. 1 NP-40 extraction of mitochondria leads to enrichment of the
or absence of cycloheximide at 100 lg/ml, at water-soluble chlor-
ATPase complex. In all, 20 lg of total mitochondrial protein and
amphenicol 50 lg/ml (Sigma, Switzerland), erythromycin at
10 lg of the 0.5% NP-40 pellet were analyzed with an antiserum
100 lg/ml, 1 mM puromycin, or a mixture of 2 lM valinomycin
directed against the ATPase complex of C. fasciculata and iodinated
and 50 lM carbonyl cyanide tri¯uoromethoxyphenylhydrazone
protein A. The antiserum recognized two components (arrows) of the
ATPase complex in T. brucei, both of which showed 3- to 5-foldenrichment in the NP-40 pellet
puromycin, a tRNA analogue that blocks prokaryotic aswell as eukaryotic translation, is added, no labeled
Mitochondria isolated by conventional hypotonic puri-
®cation procedures (Braly et al. 1974; Harris et al. 1990)
Most importantly, incorporation of radioactivity into
are not capable of synthesizing proteins in an in organ-
respiratory complexes was sensitive to erythromycin and
ello system (data not shown). We therefore used mito-
chloramphenicol, which have been shown in other sys-
chondria isolated under isotonic conditions using
tems to inhibit mitochondrial and prokaryotic transla-
nitrogen cavitation followed by Nycodenz gradients.
tion only. Antibiotic inhibition studies are the most
Mitochondria isolated by this method had previously
powerful tools for the operational de®nition of mito-
been shown to exhibit a membrane potential and to be
chondrial translation. However, previous studies have
capable of importing mitochondrial precursor proteins
led to contradictory results and have been questioned
(Hauser et al. 1996). Mitochondria were incubated un-
because the permeability of the trypanosomatid cell
der suitable buer conditions with radioactive methio-
membrane has not been addressed (Laub-Kupersztejn
nine and cysteine for 60 min at 27 °C. After incubation,
and Thirion 1974; Kleisen and Borst 1975; Spithill et al.
mitochondria were extracted with 0.5% NP-40 and the
1981). These problems can be avoided by the use of
labeled proteins were analyzed on TRIS-Tricine gels
permeabilized cells or, as in the present study, by the use
optimized to separate hydrophobic proteins (SchaÈgger
of isolated organelles. Our results fully agree with the
and von Jagow 1987). The respiratory complexes, which
study of Shu and GoÈringer (1998) and extend it further,
include practically all mitochondrially encoded proteins,
showing that radioactivity is indeed incorporated into
are relatively insoluble in 0.5% NP-40 and should
proteins. In summary, the two studies ®rmly establish
therefore be enriched in the pellet (Speijer et al. 1996).
that mitochondrial translation in trypanosomes, as in all
The immunoblot depicted in Fig. 1 shows that the
other eukaryotes, is sensitive to chloramphenicol.
ATPase complex is indeed enriched in the 0.5% NP-40
Surprisingly, choramphenicol sensitivity has not been
pellet by a factor of 3±5. During the in organello assay,
tested in a recent study on in organello translation in
radioactive methionine and cysteine is incorporated into
C. fasciculata (Tittawella 1998). A comparison of our
a subset of distinct proteins found in the NP-40 pellet.
data with these experiments is therefore problematic;
Labeling of the proteins is not altered in the presence of
nevertheless, an apparently similar labeling pattern was
cycloheximide, an inhibitor of translation in the eu-
obtained. In our study, approximately 10±12 putative
karyotic cytosol. This indicates that the observed label-
mitochondrially encoded proteins were detected. The
ing pattern was not due to cytosolic contamination or to
apparent molecular weights of the main erythromycin-
a small residual number of intact cells that might have
and chloramphenicol-sensitive labeled proteins (aster-
remained in the mitochondrial fraction. However, if
isks in Figs. 1, 2) correspond to 17.8, 22.9, and 27 kDa,
Fig. 3 Mitochondrial translation in S. cerevisiae and T. brucei arecomparable. Left panel: 20 lg of isolated yeast mitochondria was
Fig. 2 Mitochondrial translation in T. brucei is sensitive to
subjected to in organello labeling in the presence of cycloheximide, a
prokaryotic type translation inhibitors. NP-40 pellets of in organello
mixture of valinomycin and FCCP, or erythromycin and then
labeling reactions performed in the absence or presence of
analyzed on the same TRIS-Tricine gel. Right panel: same conditions
cycloheximide, chloramphenicol, erythromycin, and puromycin were
described above except that 200 lg of isolated trypanosomal
analyzed on TRIS-Tricine gel. The positions of molecular-weight
mitochondria was used in each labeling reaction and the samples
markers are indicated on the right. The three main translation
were subjected to 0.5% NP-40 extraction prior to the analysis. The
products in mitochondria are indicated by asterisks
three main translation products of T. brucei mitochondria areindicated by asterisks
respectively, which lie within the range of several pro-teins predicted to be encoded in the mitochondrion of
Acknowledgements We thank Elke Horn for her excellent techni-
T. brucei. However, the present results do not allow their
cal assistance, Rob Benne for his helpful discussions, and RolandPach for reading the manuscript. This work was supported by grant
31-46628.96 from the Swiss National Foundation and by a fel-
Finally, in organello translation in trypanosomes and
lowship awarded by the Prof. Dr. Max CloeÈtta Foundation to A. S.
in the yeast Saccharomyces cerevisiae was compared(Fig. 3). As expected, similar numbers of labeled pro-teins were found, which did not have exactly corre-
sponding sizes in the two organisms. In both species,mitochondrial translation was sensitive to erythromycin.
Beattie DS, Howton MM (1996) The presence of rotnone-sensitive
Finally, if the membrane potential was dissipated by
NADH dehyrogenase in the long slender bloodstream and the
valinomycin and FCCP, mitochondrial translation was
procyclic forms of Trypanosoma brucei brucei. Eur J Biochem241: 888±894
reduced in the yeast and abolished in T. brucei. Trans-
Benne R, Sloof P (1987) Evolution of the mitochondrial protein
port of amino acids across the inner membrane requires
synthetic machinery. Biosystems 21: 51±68
a membrane potential. It is therefore likely that the re-
Braly P, Simpson L, Kretzer F (1974) Isolation of kinetoplast-
duction in translation observed in the absence of a
mitochondrial complexes from Leishmania tarentolae. J Proto-zool 21: 782±790
membrane potential was an indirect eect due to re-
Brekk CKD, Speijer D, Dekker H, Muijsers AO, Benne R (1997)
duced uptake of amino acids. Nevertheless, such an ef-
Further evidence for the presence of mitochondrially encoded
fect is expected to be speci®c for mitochondria.
subunits in cytochrome c oxidase of the trypanosomatid Cri-
In summary, our results present for the ®rst time di-
thidia fasciculata. Biol Chem 378: 837±841
Harris ME, Moore DR, Hajduk SL (1990) Addition of uridines to
rect evidence for translation in isolated mitochondria of
edited RNAs in trypanosome mitochondria occurs indepen-
T. brucei. The observed mitochondrial protein synthesis
dently of transcription. J Biol Chem 265: 11368±11376
shows the expected properties of being resistant to
Hauser R, Pypaert M, HaÈusler T, Horn EK, Schneider A (1996)
translation inhibitors of the cytosolic type but being
In vitro import of proteins into mitochondria of Trypanosoma
sensitive to prokaryotic type translation inhibitors. Fu-
brucei and Leishmania tarentolae. J Cell Sci 109: 517±523
Kleisen CM, Borst P (1975) Are 50% of all cellular proteins syn-
ture work will focus on the identi®cation of the mito-
thesized on mitochondrial ribosomes in Crithidia luciliae? Bio-
chondrially synthesized gene products.
Laub-Kupersztejn R, Thirion J (1974) Existence of two distinct
Shu HH, GoÈringer HU (1998) Trypanosoma brucei mitochondrial
protein synthesis systems in the trypanosomatid Crithidia
ribonucleoprotein complexes which contain 12S and 9S ribo-
luciliae. Biochem Biophys Acta 340: 314±322
SchaÈgger H, Jagow G von (1987) Tricine-sodium dodecyl sulfate-
Sollner-Webb B (1996) Trypanosome RNA editing: resolved. Sci-
polyacrylamide gel electrophoresis for the separation of pro-
teins in the range from 1 to 100 kDa. Anal Biochem 166: 368±
Speijer D, et al. (1996) Puri®cation and characterization of cyto-
chrome c oxidase from the insect trypanosomatid Crithidia
Schneider A (1994) Import of RNA into mitochondria. Trends Cell
fasciculata. Mol Biochem Parasitol 79: 47±59
Spithill TW, Shimer SP, Hill GC (1981) Inhibitory eect of
Shaw JM, Campbell D, Simpson L (1989) Internal frameshifts
chloramphenicol isomers and other antibiotics on protein syn-
within the mitochondrial genes for cytochrome oxidase subunit
thesis and respiration in Trypanosoma brucei brucei. Mol Bio-
II and maxicircle unidenti®ed reading frame 3 of Leishmania
tarentolae are corrected by RNA editing: evidence for transla-
Tittawella I (1998) Protein synthesis in mitochiondria isolated from
tion of the edited cytochrome oxidase subunit II mRNA. Proc
the trypanosomatid protozoan Crithidia fasciculata. FEBS Lett
HOW TO USE THIS DIRECTORY 1,589 Mexican Corporations with sales over US $ 10’000,000 are listed in alphabetical order. Six indexes help to find them by: - Name of corporation, key words in its name, ticker symbol, usual name and name of foreign and Mexican companies related to it (pages 461-644). - North American Industry Classification System code (pages 645-- Products exported (pages
BEFORE THE ALASKA OFFICE OF ADMINISTRATIVE HEARINGS ON REFERRAL FROM THE DEPARTMENT OF ADMINISTRATION ORDER GRANTING SUMMARY ADJUDICATION S E is a retiree under the Teachers’ Retirement System (“TRS”) and is entitled to benefits of the AlaskaCare Retiree Health Plan (“Plan”). On September 26, 2007, Mr. E underwent radical retropubic prostatectomy surgery at the Mayo Clinic.
 spin (300 g) was routinely performed before loading of the Ny-codenz gradients. The absence of intact T. brucei cells and bacterialcontamination in isolated mitochondrial preparations was con-®rmed by light microscopy. In organello translations were per-formed using 400 lg of isolated mitochondria each in 100 ll oftranslation buer [20 mM TRIS-HCl (pH 7.4), 30 mM KH2PO4,0.6 M sorbitol, 5 mM succinate, 50 mM KCl, 20 mM MgSO4,12 mM creatine phosphate, creatine phosphokinase at 0.16 mg/ml,4 mM ATP, 0.5 mM GTP, 5 mM NADH, fatty-acid-free bovineserum albumin at 2.5 mg/ml, and 1 mM of all biological aminoacids except methionine and cysteine] containing 1 lM (approxi-mately 150 lCi) of a mixture of 35S-labeled methionine and cys-teine (Pro-mix, Amersham, Switzerland). The reactions wereincubated for 90 min at 25 °C, and mitochondria were reisolatedby centrifugation for 5 min at 5,200 g at 4 °C. The resulting pelletswere extracted with 100 ll of 50 mM potassium phosphate (pH7.5) containing 0.5% NP-40. Subsequently, the samples were cen-trifuged at 15,000 g for 15 min, and the pellet was solubilized underconstant mixing for 2 h at 37 °C in 50 ll of 0.28 M TRIS-HCl (pH6.8), 2.8% sodium dodecyl sulfate, 5% (v/v) glycerol, and 5% (v/v)b-mercaptoethanol. All reactions were analyzed on TRIS-Tricinegels (SchaÈgger and von Jagow 1987), which were ®rst stained withCoomassie brilliant blue to check for equal loading and then pro-cessed for ¯uorography.
spin (300 g) was routinely performed before loading of the Ny-codenz gradients. The absence of intact T. brucei cells and bacterialcontamination in isolated mitochondrial preparations was con-®rmed by light microscopy. In organello translations were per-formed using 400 lg of isolated mitochondria each in 100 ll oftranslation buer [20 mM TRIS-HCl (pH 7.4), 30 mM KH2PO4,0.6 M sorbitol, 5 mM succinate, 50 mM KCl, 20 mM MgSO4,12 mM creatine phosphate, creatine phosphokinase at 0.16 mg/ml,4 mM ATP, 0.5 mM GTP, 5 mM NADH, fatty-acid-free bovineserum albumin at 2.5 mg/ml, and 1 mM of all biological aminoacids except methionine and cysteine] containing 1 lM (approxi-mately 150 lCi) of a mixture of 35S-labeled methionine and cys-teine (Pro-mix, Amersham, Switzerland). The reactions wereincubated for 90 min at 25 °C, and mitochondria were reisolatedby centrifugation for 5 min at 5,200 g at 4 °C. The resulting pelletswere extracted with 100 ll of 50 mM potassium phosphate (pH7.5) containing 0.5% NP-40. Subsequently, the samples were cen-trifuged at 15,000 g for 15 min, and the pellet was solubilized underconstant mixing for 2 h at 37 °C in 50 ll of 0.28 M TRIS-HCl (pH6.8), 2.8% sodium dodecyl sulfate, 5% (v/v) glycerol, and 5% (v/v)b-mercaptoethanol. All reactions were analyzed on TRIS-Tricinegels (SchaÈgger and von Jagow 1987), which were ®rst stained withCoomassie brilliant blue to check for equal loading and then pro-cessed for ¯uorography.


 Fig. 3 Mitochondrial translation in S. cerevisiae and T. brucei arecomparable. Left panel: 20 lg of isolated yeast mitochondria was
Fig. 2 Mitochondrial translation in T. brucei is sensitive to
subjected to in organello labeling in the presence of cycloheximide, a
prokaryotic type translation inhibitors. NP-40 pellets of in organello
mixture of valinomycin and FCCP, or erythromycin and then
labeling reactions performed in the absence or presence of
analyzed on the same TRIS-Tricine gel. Right panel: same conditions
cycloheximide, chloramphenicol, erythromycin, and puromycin were
described above except that 200 lg of isolated trypanosomal
analyzed on TRIS-Tricine gel. The positions of molecular-weight
mitochondria was used in each labeling reaction and the samples
markers are indicated on the right. The three main translation
were subjected to 0.5% NP-40 extraction prior to the analysis. The
products in mitochondria are indicated by asterisks
three main translation products of T. brucei mitochondria areindicated by asterisks
respectively, which lie within the range of several pro-teins predicted to be encoded in the mitochondrion of
Acknowledgements We thank Elke Horn for her excellent techni-
T. brucei. However, the present results do not allow their
cal assistance, Rob Benne for his helpful discussions, and RolandPach for reading the manuscript. This work was supported by grant
31-46628.96 from the Swiss National Foundation and by a fel-
Finally, in organello translation in trypanosomes and
lowship awarded by the Prof. Dr. Max CloeÈtta Foundation to A. S.
Fig. 3 Mitochondrial translation in S. cerevisiae and T. brucei arecomparable. Left panel: 20 lg of isolated yeast mitochondria was
Fig. 2 Mitochondrial translation in T. brucei is sensitive to
subjected to in organello labeling in the presence of cycloheximide, a
prokaryotic type translation inhibitors. NP-40 pellets of in organello
mixture of valinomycin and FCCP, or erythromycin and then
labeling reactions performed in the absence or presence of
analyzed on the same TRIS-Tricine gel. Right panel: same conditions
cycloheximide, chloramphenicol, erythromycin, and puromycin were
described above except that 200 lg of isolated trypanosomal
analyzed on TRIS-Tricine gel. The positions of molecular-weight
mitochondria was used in each labeling reaction and the samples
markers are indicated on the right. The three main translation
were subjected to 0.5% NP-40 extraction prior to the analysis. The
products in mitochondria are indicated by asterisks
three main translation products of T. brucei mitochondria areindicated by asterisks
respectively, which lie within the range of several pro-teins predicted to be encoded in the mitochondrion of
Acknowledgements We thank Elke Horn for her excellent techni-
T. brucei. However, the present results do not allow their
cal assistance, Rob Benne for his helpful discussions, and RolandPach for reading the manuscript. This work was supported by grant
31-46628.96 from the Swiss National Foundation and by a fel-
Finally, in organello translation in trypanosomes and
lowship awarded by the Prof. Dr. Max CloeÈtta Foundation to A. S.