Tadalafil gehört zur Gruppe der PDE5-Hemmer und wirkt über eine hochselektive Blockade des Enzyms Phosphodiesterase Typ 5. Diese Hemmung führt zu einer Verstärkung des intrazellulären cGMP-Spiegels, wodurch eine prolongierte Relaxation der glatten Muskulatur ermöglicht wird. Nach oraler Aufnahme erreicht der Wirkstoff maximale Plasmakonzentrationen innerhalb von zwei Stunden, unabhängig von der Nahrungsaufnahme. Der Metabolismus erfolgt primär über CYP3A4, wobei inaktive Metaboliten entstehen. Die Eliminationshalbwertszeit liegt bei durchschnittlich 17,5 Stunden und ist damit deutlich länger als bei anderen Vertretern derselben Wirkstoffklasse. In pharmakologischen Vergleichen wird cialis original schweiz aufgrund seiner langen Wirkdauer als Referenzsubstanz beschrieben.
Fisica.mib.infn.it
Dynamics of green fluorescent protein mutant2in solution, on spin-coated glasses, and encapsulatedin wet silica gels
GIUSEPPE CHIRICO,1,5 FABIO CANNONE,1,5 SABRINA BERETTA,1,5ALBERTO DIASPRO,2,5 BARBARA CAMPANINI,3,5 STEFANO BETTATI,3,4,5ROBERTA RUOTOLO,3 AND ANDREA MOZZARELLI3,51Department of Physics, University of Milan-Bicocca, 20126 Milan, Italy2Department of Physics, University of Genova, 16132 Genova, Italy3Department of Biochemistry and Molecular Biology, University of Parma, 43100 Parma, Italy4Institute of Physical Sciences, University of Parma, 43100 Parma, Italy5Italian National Institute for the Physics of Matter, INFM, Italy
(RECEIVED November 13, 2001; FINAL REVISION February 8, 2002; ACCEPTED February 8, 2002)
Abstract
Single-molecule experiments are performed by investigating spectroscopic properties of molecules eitherdiffusing in and out of the observation volume or fixed in space by different immobilization procedures. Toevaluate the effect of immobilization methods on the structural and dynamic properties of proteins, a highlyfluorescent mutant of the green fluorescent protein, GFPmut2, was spectroscopically characterized in bulksolutions, dispersed on etched glasses, and encapsulated in wet, nanoporous silica gels. The emissionspectrum, the fluorescence lifetimes, the anisotropy, and the rotational correlation time of GFPmut2,encapsulated in silica gels, are very similar to those obtained in solution. This finding indicates that the gelmatrix does not alter the protein conformation and dynamics. In contrast, the fluorescence lifetimes ofGFPmut2 on glasses are two- to fourfold higher and the fluorescence anisotropy decays yield almost nophase shifts. This indicates that the interaction of the protein with the bare glass surface induces a significantstructural perturbation and severely restricts the rotational motion. Single molecules of GFPmut2 on glassesor in silica gels, identified by confocal image analysis, show a significant stability to illumination withbleaching times of the order of 90 and 60 sec, respectively. Overall, these data indicate that silica gelsrepresent an ideal matrix for following biologically relevant events at a single molecule level. Keywords: Protein immobilization; green fluorescent protein; fluorescence spectroscopy; protein dynamics; silica gels; confocal imaging
The green fluorescent protein (GFP) was discovered in the
complex cellular processes (Chalfie et al. 1994; Cubitt et al.
early 1960s (Shimomura et al. 1962), but only recently it
1995; Heim and Tsien 1996; Chalfie and Kain 1998). The
has sparked a lot of interest as a biological tool to monitor
chromophore that confers the typical green color and fluo-rescent properties to the protein is a p-hydroxybenzyli-deneimidazole, originated from an internal cyclization at
Reprint requests to: Giuseppe Chirico, Department of Physics, Univer-
residues Ser65, Tyr66, and Gly67, and 1,2 dehydrogenation
sity of Milan-Bicocca, 20126 Milan, Italy; e-mail: giberto.chirico@mib. infn.it; fax: 39-02-64482894 or Andrea Mozzarelli, Department of Bio-
of Tyr66 (Cubitt et al. 1995). The three-dimensional struc-
chemistry and Molecular Biology, University of Parma, 43100 Parma,
ture of the WT GFP and several mutants have been det-
Italy; e-mail: [email protected]; fax: 39-0521-905151.
emined (Ormo et al. 1996; Yang et al. 1996; Brejc et al. Abbreviations: GFP, green fluorescent protein; GFPmut2, GFP mutant
containing the triple substitution S65A, V68L, S72A; TPE, two-photon
1997; Palm et al. 1997; Wachter et al. 1998; Phillips 1997;
excitation; ACF, auto-correlation function; FCS, fluorescent correlation
Battistutta et al. 2000). The protein shows a -can fold con-
spectroscopy; Tris, tris(hydroxymethyl)aminomethane.
Article and publication are at http://www.proteinscience.org/cgi/doi/
␣-helix to which the chromophoric moiety is linked.
The color is completely but reversibly abolished on unfolding. Protein Science (2002), 11:1152–1161. Published by Cold Spring Harbor Laboratory Press. Copyright 2002 The Protein Society
Spectroscopic properties of immobilized GFPmut2
The spectroscopic properties of GFP have been intensively
resolved one- and two-photon fluorescence spectroscopy,
investigated (Tsien 1998 and references therein; Volkmer et al.
and fluorescence correlation spectroscopy (FCS) were per-
2000). The WT protein shows a predominant absorption band
formed on concentrated protein solutions and at single mol-
centered at 397 nm, attributed to the neutral form of the chro-
ecule level. Results clearly indicate that encapsulation of
mophore, and a lower intensity band at 470 nm, attributed to
GFP in silica gels does not perturb protein dynamics and,
the anionic form of the chromophore. The transition between
thus, is a powerful strategy for single-molecule experi-
the two species is controlled by a single ionizable residue with
a pK of ∼4.5 for the WT GFP and between 5.8 and 7.9 for
different mutants (Terry et al. 1995; Patterson et al. 1997;Haupts et al. 1998; Elsliger et al. 1999). The anionic form is a
highly fluorescent species (Tsien 1998). Independent of theexcitation wavelength, the emission band is observed at 504
nm, indicating that a proton transfer process takes place in theexcited state. Several single or multiple mutations of GFP were
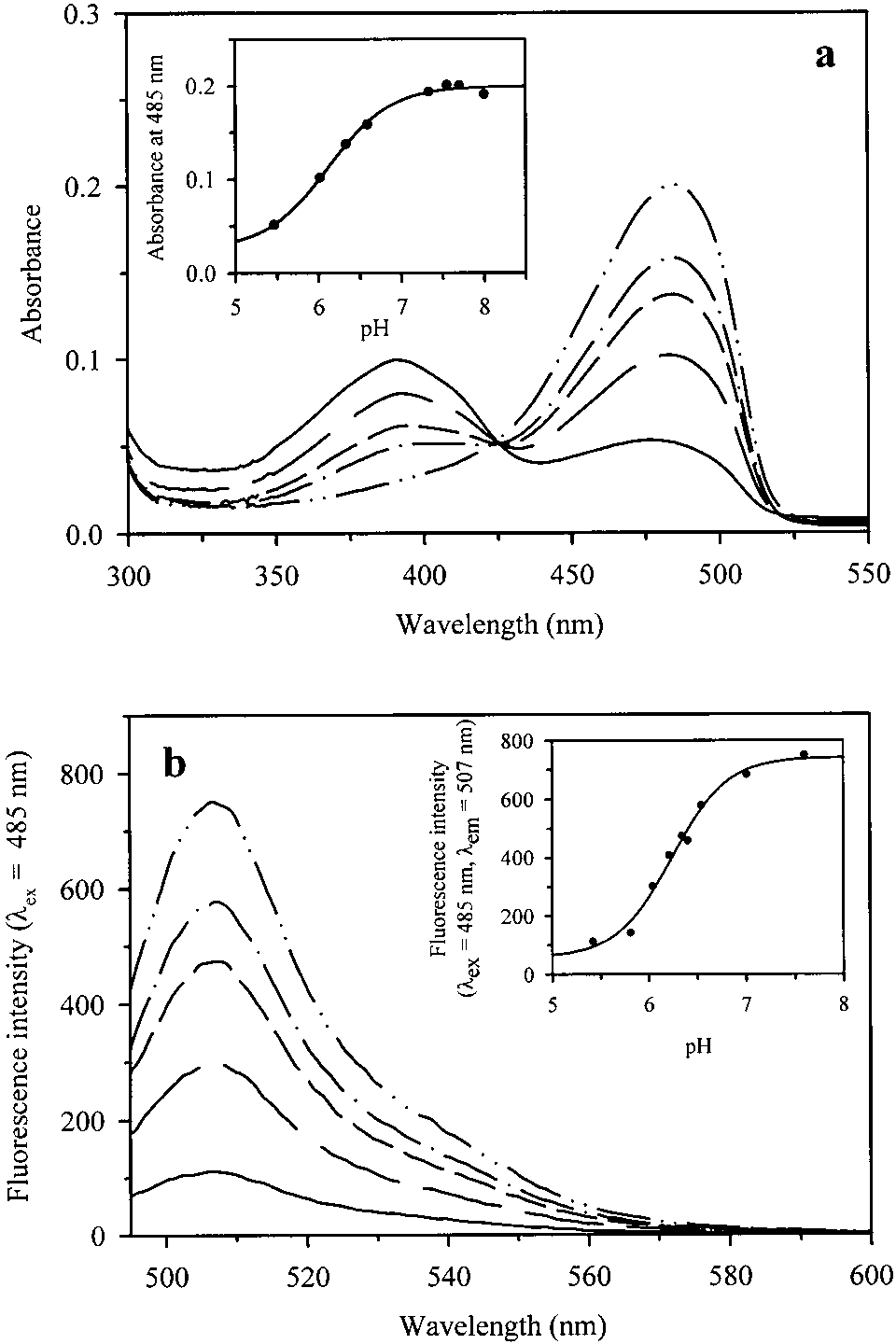
Absorbance and fluorescence spectra
obtained by random and site-directed mutagenesis to modify
the spectral properties and increase the folding efficiency. In
The absorbance spectrum of GFPmut2 shows a band cen-
particular, mutations involving Ser65 lead to the selective sta-
tered at 485 nm at alkaline pH (Fig. 1a) (Cormack et al.
bilization of the anionic form (Tsien 1998).
1996). The fluorescence emission spectrum, on excitation at
The photophysics of the fluorescent emission of WT GFP
485 nm, shows a band centered at 507 nm (Fig. 1b) (Cor-
and mutants were investigated by fluorescence up-conver-
mack et al. 1996). Similar to WT GFP (Haupts et al. 1998),
sion spectroscopy (Chattoraj et al. 1996), fluorescent cor-
absorbance and emission are pH dependent (Fig. 1a,b, in-
relation spectroscopy (Terry et al. 1995; Haupts et al.,
sets). The pH dependence was monitored between 5.5 and
1998), spectral hole-burning (Creemers et al. 1999, 2000),
8.0, as lower pH values induced protein precipitation. The
and one- and two-photon time-resolved fluorescence (Volk-
isosbestic point at 425 nm (Fig. 1a) indicates that only the
mer et al. 2000). A peculiar property of GFP, revealed by
protonated and unprotonated forms, absorbing at 388 and
single-molecule experiments on mutants immobilized in
485 nm, respectively, are in equilibrium. A single ionizable
polyacrylamide gels (Dickson et al. 1997), was the blinking
group, showing a pK of 6.13 ± 0.13 (Fig. 1a, inset) or
and switching between ionization states on light and dark
6.24 ± 0.06 (Fig. 1b, inset), controls the distribution of
chromophoric species. The fluorescence anisotropy r of
Single-molecule experiments are performed by investi-
GFPmut2, calculated from the band area, was found to be
gating molecules either diffusing in and out of the observa-
0.31 ± 0.02, independent of pH between 6.2 and 7.5 (data
tion volume or fixed in space by different immobilization
procedures (Lu et al. 1998; Kelley et al. 2001; Edman andRigler 2000; Weiss 2000; Zhuang et al. 2000a,b; Talaga et
al. 2000). Examples of the latter case are the coating of glasssurfaces with dilute chromophore solutions, the attachment
The fluorescence decays of GFPmut2 were measured for
to gold surfaces, and the entrapment in polymeric matrices,
different excitation and emission wavelengths (Table 1).
such as polyacrylamide, polymetacrylate, and agarose gels.
Decays are well described by double exponentials. The
A very promising strategy for protein encapsulation (to our
slower component ( ס 3.5 ± 0.2 ns and fractional ampli-
knowledge not yet applied in single-molecule experiments)
tude of 0.87 ± 0.03) dominates the emission and shows a
is the sol-gel technique (Brinker and Scherer 1990; Ellerby
slight but definite dependence on the emission wavelength.
et al. 1992; Brennan 1999; Bruno et al. 2001; Mozzarelli
These results are in agreement with previous studies on
and Bettati 2001, and references therein). A critical step of
enhanced green fluorescent protein (EGFP) (F64L/S65T)
single-molecule experiments on immobilized proteins is the
(Haupts et al. 1998) and GFP-S65T mutant (Volkmer et al.
evaluation of the influence of immobilization on protein
2000). The fluorescence polarization anisotropy decays,
structure and dynamics to validate the biological relevance
measured on excitation at 488 nm and emission at 535 nm
of these studies. To this purpose we have selected GFP as an
(Fig. 2), were best fitted with two components. The slower
ideal candidate because of its stability, highly fluorescent
decay corresponds to a rotational correlation time of
properties, and well-documented photophysics. In the pres-
ent study, the emission properties of the triple mutant
relaxation shows a very low anisotropy, r
Ser65Ala, Val68Leu, and Ser72Ala, called GFPmut2 (Cor-
and is characterized by a rotational time of 0.6 ± 0.15 ns.
mack et al. 1996), were characterized in bulk solutions,
Previous experiments indicated that the short rotational time
dispersed on spin-coated glasses, and encapsulated in wet,
is probably an artifact related to light scattering or instru-
porous silica gels. Confocal imaging, steady-state and time-
ment noise (Swaminathan et al. 1997). www.proteinscience.org Chirico et al. Table 1. GFPmut2 lifetimes in solution upon single photon excitation
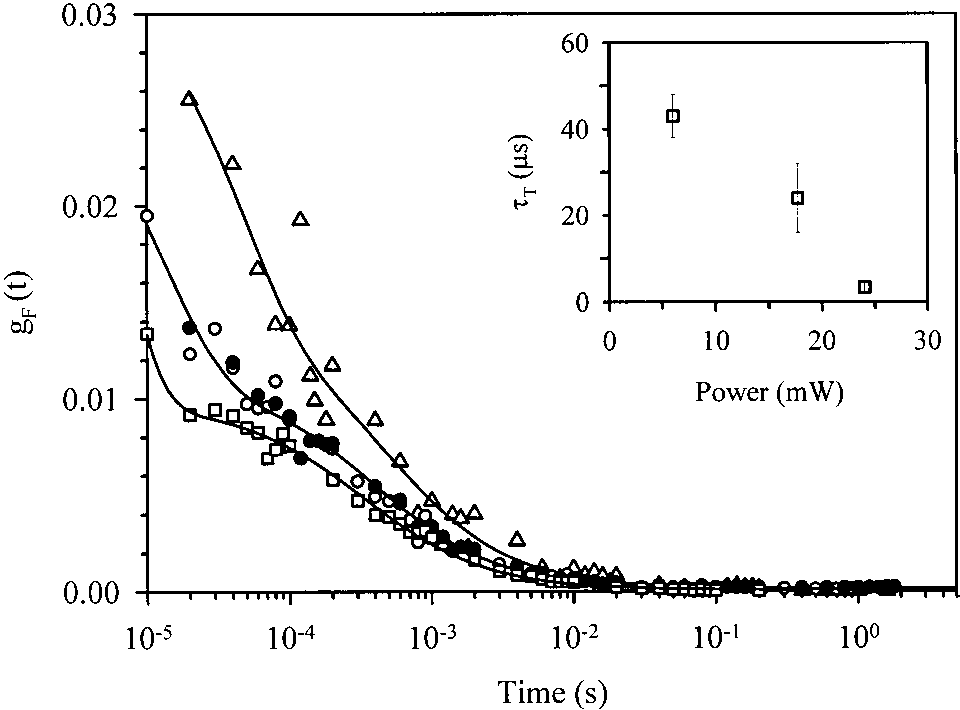
From the value of g(0) of 0.011 ± 0.002 a protein
concentration of 68 ± 20 nM was estimated, in agreementwith the expected value. From the average diffusion time of 430 ± 30 s and the measured beam waist, adiffusion coefficient D of 91 ± 6 m2/s was calculated. This value is in excellent agreement with previous dataobtained on GFP-S65T (Swaminathan et al. 1997). Theexponential relaxation in equation 3 can be related to acombination of flickering modes (reversible transitions todark states) and thermally activated delayed fluorescencefrom the triplet state of GFP (Zumbusch and Jung 2000),depending on the excitation power. Interestingly, thefraction of molecules in the dark states was approximatelyF ≅ 54 ± 10% for the two lowest excitation powers, andthe relaxation time decreased from ≅ 40 s at P ≅ 6
mW to ≅ 20 s at P ≅ 17 mW (Fig. 3, inset). Similar
results were interpreted as a light-driven flickering of
Fig. 1. (a) Absorption spectra of a solution containing GFPmut2, 10 mM
GFP (Schwille et al. 2000). At higher excitation powers,
potassium citrate, 100 mM potassium phosphate buffer, pH 5.47 (———),6.02 (— — —), 6.33 (– – – –), 6.59 (— · — · —), and 7.70 (— ·· — ·· —).
the relaxation time drops to ≅5 s, a typical value for the
(Inset) Best fit of absorbance at 485 nm to the titration of a single ionizablegroup with a pK
of 6.13 ± 0.13. (b) Fluorescence emission spectra
( ס 485 nm) of a solution containing GFPmut2, 10 mM potassium
citrate, 100 mM potassium phosphate buffer, pH 5.42 (———), 6.04 (—— —), 6.34 (– – – –), 6.54 (— · — · —), and 7.60 (— ·· — ·· —). (Inset)Best fit of fluorescence emission intensity at 507 nm to the titration of asingle ionizable group with a pK of 6.24 ± 0.06. Fluorescence correlation spectroscopy
Two-photon excitation (TPE) fluorescence correlation
spectroscopy measurements were performed using TPE at ס 800 nm on very dilute protein solutions, with an
excitation power of 6–20 mW. Representative auto-correla-tion functions (ACFs) are shown in Figure 3. For GFPmut2,the ACFs were analyzed according to a simple diffusionmodel for longer lag times plus an exponential relaxationthat corresponds to protein flickering or triplet state inter-conversion (Song et al. 1996; Haupts et al. 1998; Zumbusch
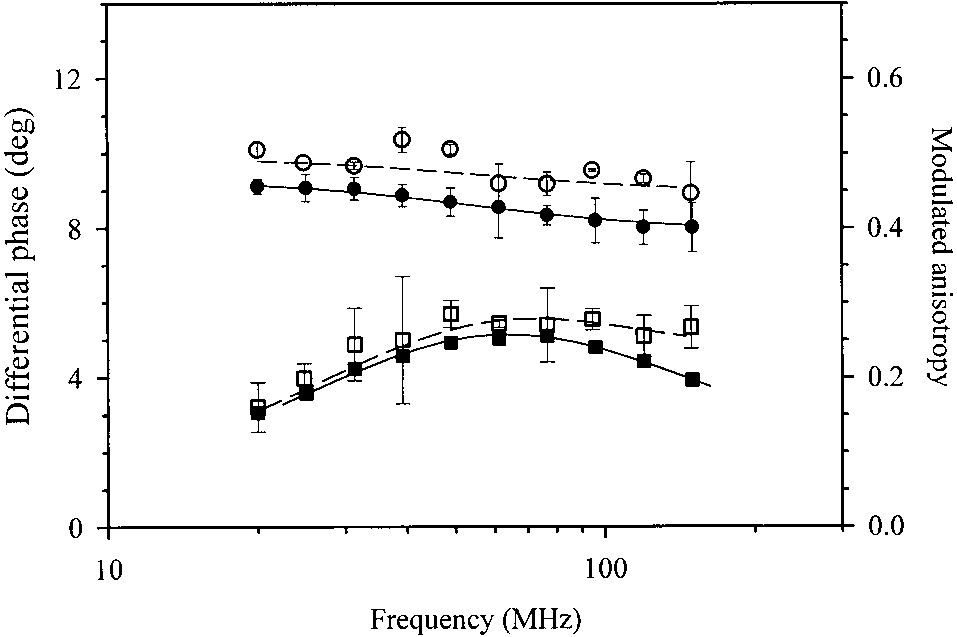
Fig. 2. Differential phase (squares) and modulated anisotropy (circles) for GFPmut2 in solution (closed symbols) and encapsulated in silica gels
(open symbols), 50 mM potassium phosphate, pH 6.7, on laser excitationat ס 488 nm. Lines represent the least-squares best fit with a two-
Protein Science, vol. 11 Spectroscopic properties of immobilized GFPmut2
tensity, shown by individual molecules (Fig. 6), are notcaused by different orientation of the molecular dipole mo-ments because the exciting light is unpolarized. In somecases, during the bright phase, a switching on/off behavioris observed. This phenomenon, known as blinking (Dicksonet al. 1997), is related to internal photodynamics of indi-vidual GFP molecules. The present acquisition time of oursetup is 229 ms per image; the time interval between con-secutive images is 458 ms, likely preventing a detailed ob-servation of blinking that is expected (for excitation inten-sity ≅700 kW/cm2) to be much faster than our resolutiontime. In fact, the on/off time varies over a wide range de-pending on the mutant GFP and the light intensity. Forexample, for EGFP it was found that <T
> ≅ 2 sec for an excitation intensity ≅14 kW/cm2
(Garcia-Parajo et al. 2000), which is considerably lower
Fig. 3. Autocorrelation functions of the emission of a solution containing
than the present value (700 kW/cm2). The WT GFP eva-
GFPmut2 (concentration of the order of 0.1 M), 50 mM phosphate buffer,
nescent wave fluorescence microscopy measurements at an
pH 7.6, on two-photon excitation (TPE) at ס 800 nm. Lines represent
the fit with a diffusion component for long lag times and an exponentialrelaxation for short lag times. Symbols refer to different excitation powers:24 mW (triangles), 17 mW (closed and open circles), and 6 mW (squares). Inset: triplet relaxation time obtained from the exponential fit of the shortlag time component of the auto-correlation functions (ACFs) as a functionof the excitation power.
thermally activated delayed fluorescence (Zumbusch andJung 2000; Schwille et al. 2000).
The representative image of glasses spin coated with 1–2
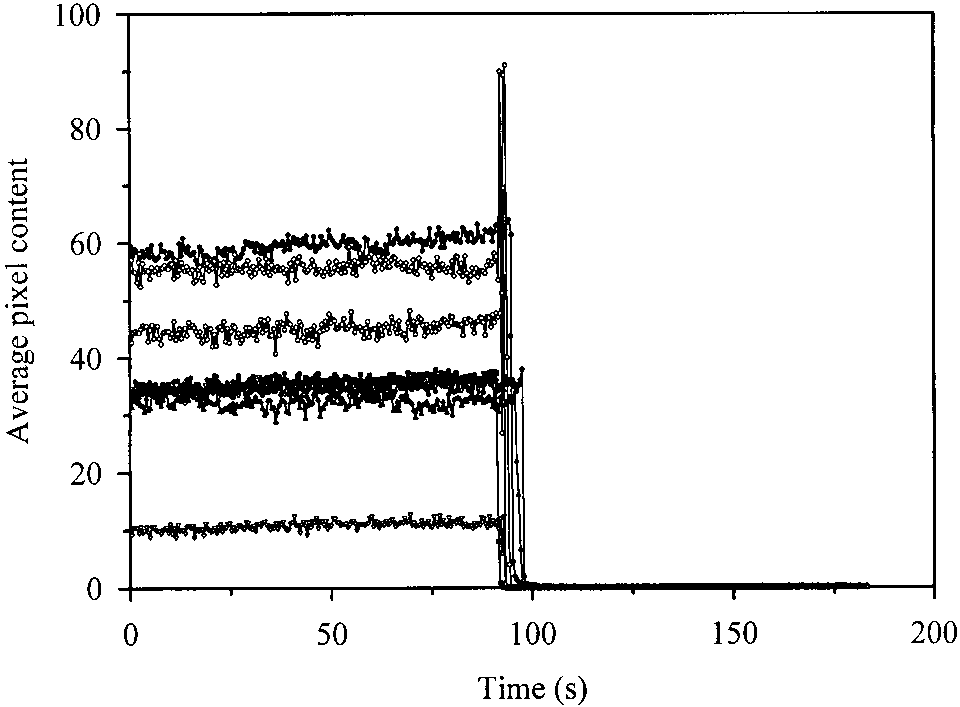
M GFPmut2 (Fig. 4a), obtained using the confocal micro-scope, showed well-defined fluorescent spots over a scat-tering background. The distribution of spot intensity, evalu-ated for each pixel (Fig. 5a), and the average pixel intensity(Fig. 5b) indicate that the dimmer spots contain singleGFPmut2 molecules, whereas more intense spots contain upto four molecules. For the single molecule spots the fluo-rescence intensity was constant with time, before a suddenincrease and a successive drop to the background level (Fig. 6). The duration of the bright phase, T
as determined from 10 spots, is 94 ± 3 sec, and decreaseslinearly with the excitation power (data not shown). Thisfinding indicates that the drop of fluorescence emission maybe attributable to a thermally induced local rearrangementof the fluorophore pocket. We do not have an explanationfor the sudden signal increase observed just before thebleaching. The background level was ≅0.03 a.u., which is
Fig. 4. Typical fluorescence image of GFPmut2, spin coated on etched
about three orders of magnitude lower than the fluorescence
glasses (a), and encapsulated in silica gel (b). The view field is 10 × 10 mand the residence time is 9 . Fluorescence imaging was performed by
output from single GFP molecules, ≅40 a.u. This high signal
confocal microscopy (see Materials and Methods) on laser excitation at
to background ratio is particularly important in single-mol-
488 nm. Protein concentration was of the order of 1 M in 50 mM potas-
ecule experiments. The different levels of fluorescence in-
www.proteinscience.org Chirico et al.
excitation power of ≅10 mW (corresponding presumably toan excitation intensity much lower than 14 kW/cm2) indi-cated an even longer <T
> ≅ 8 sec (Pierce et al. 1997).
For fluorescence lifetime measurements on etched glass
slides, solutions containing higher protein concentrationswith respect to imaging experiments were spread by spincoating. The lifetimes were measured at ס 80 MHz, using1 M rhodamine 6G in ethanol as a reference. The Ti:Sapphlaser beam was passed through the epi-fluorescence port onthe sample at a power ≅24 mW for TPE at ס 800 nm. For
Fig. 6. Kinetics of fluorescence emission ( ס 488 nm) from 10 single
GFPmut2 molecules spin-coated on etched glasses. Collection time: 458ms. Excitation power: ≅1 mW.
100 nM fluoresceine solutions spread on glasses, it wasfound that
value of lifetime was ס 4.4 ± 0.3 ns, very close to thesolution value at high pH, ≅4.0 ns. The lifetimes for ≅1 MGPFmut2,
ס 6 ± 0.7 ns and ס 12 ± 2.5 ns (data not shown).
We could not obtain measurements with sufficient accuracyfor larger modulation frequencies because of the low fluo-rescence rate of GFPmut2. GFPmut2 encapsulated in silica gelsFluorescence spectra and fluorescencesteady-state anisotropy
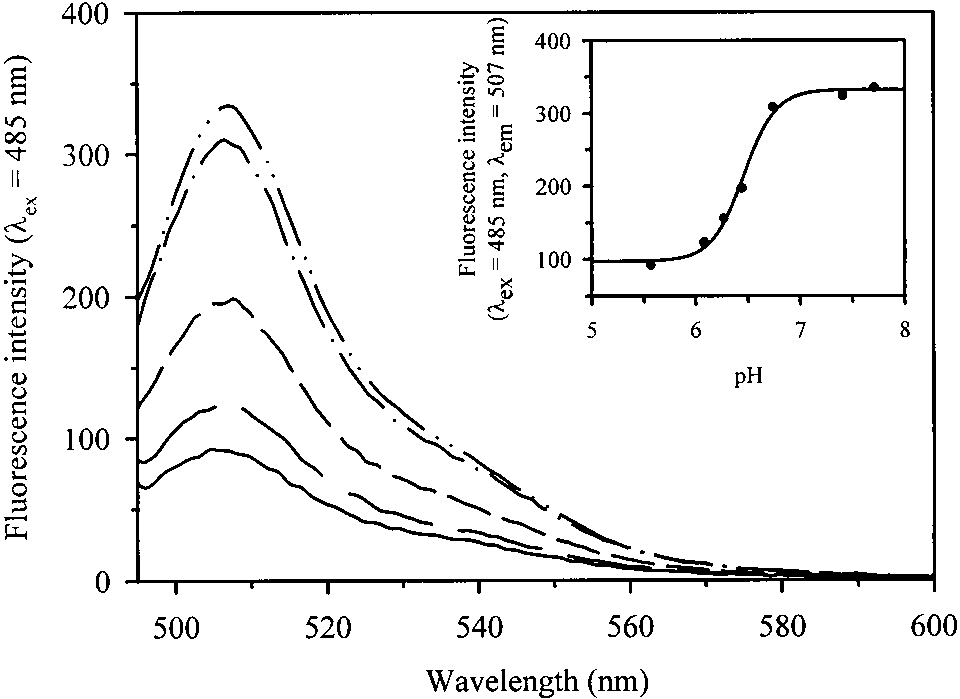
On excitation at 485 nm, the fluorescence emission spec-
trum of GFPmut2, encapsulated in wet porous silica gels,showed a peak centered at 507 nm (Fig. 7), as in solution(Fig. 1b). The pH dependence is controlled by an ionizableresidue with pK of 6.46 ± 0.03 (Fig. 7, inset), a value
slightly higher than in solution (Fig. 1b, inset). The fluores-cence anisotropy was found to be 0.38 ± 0.01.
Gels for fluorescence imaging were prepared by spread-
Fig. 5. (a) Distribution of spot average fluorescence emission of GFPmut2
ing a drop of GFPmut2-sol mixture between two glass slides
evaluated by averaging on a square of 441 pixels around the spots for 10
before gel formation and sealing to avoid drying. Imaging
similar images of protein on glass and for six images of protein in silicagels. Open and closed squares represent the histogram of the number of
performed on GFPmut2 gels with the confocal microscope
fluorescent spots with a given average fluorescence for GFPmut2 in silica
revealed bright spots on top of a background signal (Fig.
gels and on glass, respectively. The solid line is a multigaussian fit of the
4b). The distribution of the spot intensity, evaluated for each
data. (b) The mean values of the gaussian fits reported in panel (a) are
pixel (Fig. 5a), and the average pixel intensity (Fig. 5b)
plotted as a function of the aggregation order to determine the average
allow identification of spots originated from single mol-
fluorescence intensity per spot. The open and closed circles refer to data forGFPmut2 in silica gels and on glass, respectively. The solid line is a linear
ecules of GFPmut2 (Chirico et al. 2001). The less bright
spots show a stable fluorescence emission with T
Protein Science, vol. 11 Spectroscopic properties of immobilized GFPmut2
silica gels—were investigated to evaluate the effect of dif-ferent immobilization procedures on the dynamic propertiesof the protein. This information is instrumental to establishthe biological and biotechnological relevance of spectro-scopic studies on single immobilized molecules. Single-molecule experiments can be performed also in solution byon-the-fly experiments (Deschenes et al. 2001; Weiss 1999,2000; Volkmer et al. 2000; Haupts et al. 1998) probing theconformational ensemble of molecules while diffusing inthe excitation volume. However, experiments on immobi-lized molecules offer the possibility to monitor a widerrange of dynamical processes and their kinetics. Experi-ments have been performed on proteins either fixed onetched glasses by chemi- or physi-adsorption (Talaga et al. 2000) or entrapped in polymeric matrices as polyacrylamide(Dickson et al. 1997) and agarose gels (Lu et al. 1998). We
Fig. 7. pH-dependence of fluorescence emission spectra ( ס 485 nm)
are not aware of any single-molecule study on proteins en-
of GFPmut2 encapsulated in silica gels. Gels were soaked in a solutioncontaining 10 mM potassium citrate, 100 mM potassium phosphate buffer,
pH 5.57 (———), 6.08 (— — —), 6.44 (– – – –), 6.74 (— · — · —), and
We first investigated the static and dynamic properties of
7.71 (— ·· — ·· —). (Inset) Best fit of fluorescence emission intensity at
GFPmut2 in solution. The absorption and emission spectra
507 nm to the titration of a single ionizable group with pK of 6.46 ± 0.03.
of GFPmut2 show bands slightly blue-shifted with respectto EGFP (Patterson et al. 1997), and their dependence on pH
62.4 ± 3.9 sec (Fig. 8). It is worth noting that (1) the fluo-
is controlled by a single ionizable residue with a pK similar
rescence level of different single GFPmut2 molecules
to that observed for EGFP (Haupts et al. 1998).
within the silica gels is remarkably similar, in contrast with
The overall size of the protein was determined both from
GFPmut2 on glass slides (Fig. 5); (2) no sudden increase of
the translational diffusion coefficient measured by FCS and
fluorescence was observed before the signal drop; and (3)
the rotational diffusion time provided by the analysis of
the intensity of fluorescence arising from a single molecule
fluorescent polarization anisotropy data. From FCS mea-
is approximately twice the background signal in the gel. As
surements we determined D ס 92 ± 6 m2/s, which corre-
in the case of GFPmut2 spin-coated glasses, in some cases,
sponds to a hydration radius R ס 2.4 ± 0.17 nm, if a
switching on-off behavior of the fluorescence signal was
simple spherical symmetry is assumed. The rotational time
in solution, ס 12.5 ± 0.3 ns, corresponds to a rotational
Time-resolved fluorescenceLifetime measurements were performed on GFPmut2
gels with single photon excitation ( ס 488 nm) at 12modulation frequencies in the range of 20–150 MHz. Fromthe analysis of the data, a lifetime of 3.5 ± 0.2 ns with
f ס 0.82 ± 0.04 and a lifetime of 1.1 ± 0.2 ns with
f ס 0.18 ± 0.03 were calculated, in very close agreement
with solution data. The fluorescence polarization anisotropywas measured on silica gels at higher GFPmut2 concentra-tion. The decays are reported in Figure 2. Although theuncertainties on the gel data are larger because of the lowerconcentration and the residual scattering from the gel, thefluorescence decays of GFPmut2 gels are remarkably simi-lar to those observed in solution. The analysis of the dataindicate two FPA rotational times,
anisotropy r ס 0.34 ± 0.06 and a fast relaxation time
ס 0.3 ± 0.1 ns with anisotropy r ס 0.05 ± 0.002. Fig. 8. Kinetics of fluorescence emission ( ס 488 nm) from single Discussion
green fluorescent protein (GFP) molecules encapsulated in silica gels. Gelswere soaked in a solution containing 10 mM potassium citrate and 100 mM
The fluorescence properties of the mutant GFPmut2 in three
potassium phosphate buffer, pH 7.6. Collection time: 458 ms. Excitation
different environments—solution, etched glasses, and wet
www.proteinscience.org Chirico et al.
diffusion coefficient ⌰ ס13.3 ± 0.3 MHz. Assuming a
measurements of fluorescence emission of S65T GFP, im-
spherical symmetry for the protein, an average radius
mobilized in polyacrylamide gels, indicate that the on time
R⌰ ס 2.3 ± 0.02 nm was obtained, in excellent agreement
dramatically depends on light intensity and polarization
with the hydration radius and with the values reported for
(Garcia-Parajo et al. 2000). Because the excitation light
WTGFP (Terry et al. 1995), EGFP (Haupts et al. 1998), and
used in our experiments was unpolarized, the presence of
partially unfolded molecules on the coated glasses seems to
The steady-state fluorescence properties of GFPmut2, en-
be a more likely explanation. This finding calls for a cau-
capsulated in silica gels, appear almost indistinguishable
tious interpretation of single-molecule experiments per-
from those obtained in solution. This implies that the gel
formed on biomolecules directly immobilized on glasses. In
matrix does not alter the protein conformation. Regard-
such studies, which were aimed to investigate catalysis and
ing the dynamic properties, the fluorescence lifetimes of
folding of RNA (Zhuang et al. 2000) and folding and un-
GFPmut2, encapsulated in silica gels, are very similar to the
folding of two-stranded coiled-coil peptides (Talaga et al.
values obtained in solution. In contrast, the fluorescence
2000), severe controls were performed.
lifetimes of GFPmut2 on glasses are two- to fourfold higher.
In the case of immobilization of proteins in silica gels,
This indicates that the interaction with the bare glass surface
functional properties are not significantly perturbed, as
induces a significant perturbation of protein structure and
proven for several enzymes (Bettati and Mozzarelli 2001,
dynamics that is not observed when the protein is entrapped
and references therein) and hemoglobin (Bettati and Moz-
in silica gels. Moreover, the rotational correlation time of
zarelli 1997; Bruno et al. 2001; Abbruzzetti et al. 2001b).
the protein in silica gels, ס 15 ± 2 ns, is only slightly
The influence of immobilization on dynamic properties may
higher than the value observed in solution, ס 12.5 ± 0.3
vary among different proteins depending on constraints
ns, indicating that the constraints imposed by the gel matrix
caused by specific interactions with the negatively charged
on protein rotation are limited. Similar experiments on
silica matrix. In the case of albumin labeled with acrylodan,
glasses yield almost no phase shifts, although with large
time-resolved anisotropy measurements indicated a small
uncertainties (data not shown), indicating that the rotational
decrease in the global motion of the protein and an unre-
motion of the protein physi-adsorbed on the glasses is se-
stricted local motion of the probe with respect to the protein
verely restricted, as reported for other molecules embedded
in solution (Jordan et al. 1995). In the case of silica gel-
in various types of gels (Deschenes et al. 2001).
encapsulated myoglobin, rotational diffusion was signifi-
To investigate biologically relevant processes on single
cantly impeded (Gottfried et al. 1999), and the amplitude of
molecules, such as protein folding and unfolding and en-
the carbon monoxide geminate rebinding was increased
zyme catalysis, a critical requirement is the photostability of
(Abbruzzetti et al. 2001a). Interestingly, myoglobin shows a
the excited chromophore. It is well known that Trp residues
positively charged surface, whereas GFP is negatively
are very rapidly bleached by the intense light pulse used in
charged. This finding indicates that silica gel protein encap-
single-molecule experiments and cannot be used as a probe
sulation is a well-suited method for single-molecule experi-
(Bent and Hayon 1975). The time course of the fluorescence
ments, but some caution should still be exerted.
intensity of single GFPmut2 molecules immobilized onglasses and in silica gels indicates that the fluorescencesignal arising from individual GFPmut2 molecules is easily
Materials and methods
resolved from the background. The signal/background ratiois lower in silica gels than in the case of GFPmut2 spread on
GFPmut2 expression and purification
etched glasses. Another important feature is that T
time after which the fluorescence definitely drops to the
GFPmut2 gene, cloned in a pKEN1 vector (Ezaz-Nikpay et al. 1994), was kindly provided by Dr. Brendan P. Cormack (Depart-
background level, is remarkably long for both the protein on
ment of Microbiology and Immunology, Stanford University
glasses and in silica gels and allows one to monitor bio-
School of Medicine, Stanford, CA). GFPmut2 is a GFP mutant
chemically relevant processes. Moreover, the constant fluo-
containing a triple substitution, S65A, V68L, and S72A, confer-
rescence intensity of GFPmut2 in silica gels, compared with
ring enhanced fluorescence emission and high yield of protein
the variability observed on glasses (Figs. 6, 8), provides
caused by a more efficient folding at 37°C with respect to WT(Cormack et al. 1996). For expression of GFPmut2, Escherichia
evidence for an unperturbed native conformation of the pro-
coli strain XL1-Blue was used and bacteria were grown in 2xYT
tein in silica gels. On the contrary, the different fluorescence
medium (16 g tryptone, 10 g yeast extract, 5 g NaCl per liter). A
intensities, observed for GFPmut2 on glasses (Fig. 6), might
single E. coli colony containing the recombinant construct was
be explained by either different or fixed orientations of the
inoculated in 10 mL of 2xYT medium supplemented with ampi-
molecules with respect to the excitation light or a distribu-
cillin (100 g/mL) and tetracycline (50 g/mL) and incubatedovernight at 37°C on a rotating shaker. One liter of 2xYT medium
tion of folded and partially unfolded proteins, the unfolded
containing the antibiotics was inoculated with the starter culture
GFP having completely lost the green fluorescence (Tsien
and shaken at 37°C until the cells reached the mid-log phase of
1998). Confocal and near-field scanning optical microscopy
ס 0.6). Expression of GFPmut2 was induced by
Protein Science, vol. 11 Spectroscopic properties of immobilized GFPmut2
adding isopropyl--D-galactopyranoside to a final concentration
Å in diameter) showed a typical pore structure with a pore size of
of 1 mM and incubating the culture at 37°C on a rotating shaker for
∼30–40 and 20 Å, respectively (Abbruzzetti et al. 2001b).
2.5 h. Cells were harvested by centrifugation and resuspended in50 mM Tris/HCl buffer, pH 8.0, containing 1 mM EDTA, 1 mM
-mercaptoethanol, 0.1 M NaCl, 0.5 mM phenylmethylsulphonyl-
fluoride, 0.5 mM benzamidine, 1M leupeptin, and 1 M pep-statin. Cells were sonicated until the fluorescence emission of the
The optical setup for the single-photon imaging experiments is
supernatant, on excitation at 485 nm, was constant. The crude
based on an inverted microscope (TE300, Nikon), a Nikon
extract was centrifuged at 14,000g for 45 min and streptomycin
PCM2000 scanning head, and an air-cooled Argon laser with ex-
sulfate was added to the supernatant to a final concentration of
citation wavelength at 488 nm. The laser beam is sent to the
about 3.3% (w/v). The solution was kept for 30 min at 4°C and
entrance pupil of a Nikon objective (N.A. ס 1.4, Plan Apochro-
then centrifuged for 45 min at 14,000g. The pellet was discarded
mat DICH 100X oil, working distance 0.19 mm, focal length 2
and ammonium sulfate was added to the supernatant to 40% satu-
mm) by the scanning lens. The fluorescence signal, collected by
ration. The solution was then centrifuged at 14,000g for 15 min
the same objective and selected by a HQ535–50 filter (Chroma
and the pellet was again discarded. Ammonium sulfate was added
Inc.), is fed to a single-mode fiber connected to a R928 photomul-
to the supernatant to 70% saturation. The solution was centrifuged
tiplier (Hamamatsu) in the PCM2000 controller.
at 14,000g for 15 min. The pellet was redissolved in a minimum
Fluorescent molecules, either spin coated on etched glasses or
volume of 50 mM Tris/HCl buffer, pH 8.0, containing 1 mM
encapsulated in silica gels, were imaged by the Nikon EZ-2000
EDTA and 1 mM dithiothreitol (buffer A). The solution containing
software interfaced to the PCM2000 scanning head (Diaspro et al.
GFPmut2 was dialyzed overnight against buffer A containing 0.5
1999a). For the confocal setup the resolutions are 0.19 m in the
M NaCl. Protein solution was concentrated to about 2 mL by
plane and 0.6 m in the axial direction (Diaspro et al. 1999a). The
ultrafiltration and loaded onto a size exclusion column (G-75 fine,
acquisition of the images (512 × 512 pixels) with a residence time
Amersham Pharmacia Biotech; bed volume ס 200 mL) equili-
of about 9 s per pixel takes 2.3 sec. The view field is in the range
brated with buffer A containing 0.5 M NaCl. The column was
35–140 m and the excitation power is usually about 13 mW,
eluted at a constant flow of 80 L/min. Fractions containing
which corresponds to a light intensity of 700 kW/cm2.
GFPmut2 were collected and the purity of the sample was evalu-ated by SDS-PAGE. Fractions were pooled on the basis of the
Fluorescence kinetics of individual spots
degree of purification of GFPmut2 and concentrated to about 100
M. An 85% pure protein solution was stored at −80° C and used
A wide field (80 × 80 m2) image was collected before the time
course acquisition and compared with a 3D scanning performedright after the kinetics to ensure that the disappearance of thefluorescence was not caused by a shift of the focus plane. For this
application, 160 × 160 pixels images were acquired on single spots(15 × 15 m2 field) with ≅9 s residence time. The acquisition
GFPmut2 from stock solutions was diluted in phosphate-contain-
time is 229 ms per image and the time interval between consecu-
ing buffers. All experiments were performed at room temperature.
tive images is 458 ms. The fluorescence intensity of each spot wascomputed by summing the pixel content in a circular area aroundeach spot and by normalizing for the number of pixels in this area,
which has a typical diameter of 10 pixels, corresponding to ≅0.9
m. The computation of the spot intensity was performed with a
The glass slides were first soaked in a solution containing 1%
home-coded MatLab (Mathworks, Inc.) program, which allows
sodium dodecyl-sulfate for 24 h, then in a methanol solution satu-
identification of a single spot on a time series of images and
rated with NaOH for 2 h. To remove residual traces of NaOH, the
computation of the spot intensity as a function of the acquisition
slides were first soaked in 0.1% HCl solutions for 2 h, then in a
diluted chromic solution for 2 h, and, finally, rinsed extensivelywith Milli-Q water (Millipore, Inc.). After this procedure, glasseswere stored for a few hours in ethanol. After rinsing thoroughly
with Milli-Q water and drying under a filtered nitrogen flux,
Steady-state fluorescence spectra were acquired with a Perkin-
sample solutions were spread on the glass slides by spin coating.
Elmer LB-50 spectrofluorometer. The fluorescence anisotropy Gvalues for GFP in solution and encapsulated in silica gels are1.38 ± 0.05 and 1.33 ± 0.03, respectively.
Encapsulation of GFPmut2 in silica gels was performed according
Fluorescence lifetime and anisotropy
to Bettati and Mozzarelli (1997). The stock protein solution wasdiluted 50- to 100-fold in 10 mM citrate, 100 mM phosphate
For time-resolved fluorescence measurements, the emitted light
buffer, pH 7.5. Sixty-seven L of the resulting solution were
was detected through the front port of the microscope by a R928
mixed with 100 L sol, prepared from tetramethylorthosilicate,
photomultiplier tube (Hamamatsu). The gain was modulated by
water, hydrochloric acid, and phosphate buffer. On gelation, silica
biasing the second dynode stage at a radio frequency. The cross-
gels were covered with the same buffer solution and stored at 4°C
correlation frequency was 36 Hz and the modulation frequencies
for at least 12 h before use. Gel pore size is less than ∼20 Å
were provided by a master radio frequency synthesizer (Marconi
because GFPmut2 molecules, characterized by an average diam-
Instruments, mod. 2023A) that biased a Pockels cell, which,
eter of about 40 Å, did not leach. Transmission electron micros-
coupled to a polarizer beamsplitter, modulated the amplitude of the
copy measurements of silica gels with and without hemoglobin (60
laser light intensity (Argon Laser, 2025) at frequencies in the range
www.proteinscience.org Chirico et al.
of 20–150 MHz. The photomultiplier signal was fed to an ISS
squares fitting program based on the Marquardt algorithm (Bev-
lock-in amplifier board (ISS) for the computation of the polarized
ington 1992) and using the routine MRQMIN (Press et al. 1993)
modulation ratios and phase differences of the fluorescence light
for fitting. From the analysis of the ACFs, the relaxation time, ,
with respect to the excitation laser beam. The synchronization was
and the zero lag-time term, g (0), were determined. Measurements
performed by sending the signal of a second photomultiplier, mea-
of the fluorescence fluctuations of fluorescein solutions at various
suring the intensity of a small fraction of the excitation laser beam,
concentrations (Chirico et al. 2000) provide an estimate of the laser
to the ISS board as a frequency reference.
beam waist, w , assuming a translational diffusion coefficient D
The fluorescence lifetime measurements were performed with
of 280 m2/s at 22°C (Rigler et al. 1993). The value of the exci-
the polarizer at the magic angle ( ס 54.7°). The lifetime refer-
tation volume determined for the present setup is V
ence was either an alkaline solution of fluorescein at p⌯ ≅ 8, char-
0.04 fL at ס 770 nm (Chirico et al. 2000).
acterized by a lifetime of 4.05 ns (Tsien and Wagonner 1995) orrhodamine 6G in ethanol, characterized by a lifetime of 3.89 ns(Thompson and Gratton 1988). The polarization of the fluores-
Acknowledgments
cence was selected by a Glan-Thompson polarizer (extinction ratio<10–6) and the fluorescence polarization anisotropy measurements
This work was supported in part by grants from the Italian Min-
were performed by rotating the polarizer between the directions
istry of Instruction, University and Research (PRIN2001 to A.M.)
parallel and perpendicular to that of the excitation light. The dif-
and the National Institute for the Physics of Matter (PAIS
ferential phase shifts and the polarized modulation ratios were
provided directly by the ISS acquisition program. The G factor was
The publication costs of this article were defrayed in part by
measured directly on the TE300 microscope and found to be very
payment of page charges. This article must therefore be hereby
close to unity, G ס 1.000 ± 0.005.
marked “advertisement” in accordance with 18 USC section 1734solely to indicate this fact. TPE fluorescence correlation spectroscopyReferences
The optical setup for the TPE experiments is based on the sameinverted microscope used for fluorescence imaging and a mode-
Abbruzzetti, S., Viappiani, C., Bruno, S., and Mozzarelli, A. 2001a. Enhanced
geminate ligand rebinding upon photo-dissociation of silica gels-embedded
locked Ti:sapphire laser (Tsunami 3960, Spectra Physics; pulses
myoglobin-CO. Chem. Phys. Lett. 346: 430–436.
width of about 100 fs, repetition frequency of 80 MHz). A portion
Abbruzzetti, S., Viappiani, C., Bruno, S., Bettati, S., Bonaccio, M., and Moz-
of the laser beam is sent to the entrance pupil of the objective by
zarelli, A. 2001b. Functional characterization of heme proteins encapsulated
the scanning lens. The laser power at the object plane can be
in wet nanoporous silica gels. J. Nanosci. Nanotech. 1: 407–415.
adjusted by neutral filters. Only about 30% of the laser light en-
Battistutta, R., Negro A., and Zanotti, G. 2000. Crystal structure and refolding
properties of the mutant F99S/M153T/V163A of the green fluorescent pro-
tering the scanning head excites the sample because of losses and
tein. Proteins 41: 429–437.
absorption in the confocal head and the objective. The excitation
Bent, D.V. and Hayon, E. 1975. Excited state chemistry of aromatic amino acids
power was typically 5 mW on the sample, corresponding to 900
and related peptides. III. Tryptophan. J. Am. Chem. Soc. 97: 2612–2619.
kW/cm2. The fluorescence signal was collected by the same ob-
Berland, K.M., So, P.T.C., and Gratton, E. 1995. Two-photon fluorescence
jective and selected by a filter (HQ535–50, Chroma Inc.). The
correlation spectroscopy: Method and application to the intracellular envi-
point spread function of the TPE corresponds to a plane resolution
ronment. Biophys. J. 68: 694–701.
Bettati, S. and Mozzarelli, A. 1997. T state hemoglobin binds oxygen nonco-
of ≅0.22 m and an axial resolution of ≅0.6 m (Diaspro et al.
operatively with allosteric effects of protons, inositol hexaphosphate and
1999b). For measurements of the fluorescence fluctuations, the
chloride. J. Biol. Chem. 272: 32050–32055.
photon counts are detected through the bottom port of the micro-
Bevington, P.R. 1992. Data reduction and error analysis for the physical sci-
scope by an avalanche photodiode detector (SPCM-AQ-151,
ences. McGraw Hill, NY, ch. 11, p. 204
EG&G). The output signal is fed to a correlator board (ISS) with
Brejc, K., Sixma, T.K., Kitts, P.A., Kain, S.R., Tsien, R.Y., Ormo, M., and
a sampling time of 50 s. The ACF of the fluorescence signal F(t),
Remington, S.J. 1997. Structural basis for dual excitation and photoisom-erization of Aequorea victoria green fluorescent protein. Proc. Natl. Acad.
defined as g (t) ס <␦F(t)␦F(0)>/<F(0)2, can be well approximated
Sci. 94: 2306–2311.
(Berland et al. 1995) for the TPE experiments using the equation:
Brennan, J.D. 1999. Using intrinsic fluorescence to investigate proteins en-
trapped in sol-gel derived materials. Appl. Spectrosc. 53: 106A–121A.
Brinker, C.J. and Scherer, G.W. 1990. Sol-Gel science: The physics and chem-
exp͑− tր ͒ͬg ͑0͒͑1 + tր͒−1
istry of Sol-Gel processing, Academic Press, Boston, MA.
Bruno S., Bonaccio, M., Bettati, S., Rivetti, C., Viappiani, C., Abbruzzetti, S.,
and Mozzarelli, A. 2001. High and low oxygen affinity conformations of T
The zero lag time term is related to the average number concen-
state hemoglobin. Protein Sci. 10: 2401–2410.
Chalfie, M., Tu, Y., Euskirchen, G., Ward, W.W., and Prasher, D.C. 1994.
Green fluorescent protein as a marker for gene expression. Science 263: 802–805.
Chalfie, M. and Kain, S. (eds.) 1998. Green fluorescent protein: Properties,applications and protocols. Wiley-Liss, N.Y.
Chattoraj, M., King, B.A., Bublitz, G.U., and Boxer, S.G. 1996. Ultra-fast
excited state dynamics in green fluorescent protein: Multiple states and
where the term ס 0.076 is determined by the laser beam shape
proton transfer. Proc. Natl. Acad. Sci. 93: 8362–8367.
at the objective focus, the volume of the excitation profile is
Chirico, G., Olivini, F., and Beretta, S. 2000. Fluorescence excitation volume in
related to the laser beam waist, w , by the relationship
two-photon microscopy by autocorrelation spectroscopy and photon count-
ס w 4/ (Berland et al. 1995), and B indicates the con-
ing histogram. Applied Spectrosc. 54: 1084–1090.
Chirico, G., Cannone, F., Beretta, S., Baldini, G., and Diaspro, A. 2001. Single
tribution of an uncorrelated background to the detected signal. The
molecules studies by means of the two-photon fluorescence distribution.
relaxation time is related to the translational diffusion coefficient
Microsc. Res. Tech. 55: 359–364.
D of the fluorophore, as ( 1.15w 2/(8D ) (Chirico et al. 2000).
Cormack, B.C., Valdivia, R.H., and Falkow, S. 1996. FACS-optimized mutants
The analysis of the ACFs was performed by a home-coded least
of green fluorescent protein (GFP). Gene 173: 33–38. Protein Science, vol. 11 Spectroscopic properties of immobilized GFPmut2
Creemers, T.M.H., Lock, A.J., Subramaniam, V., Jovin, T.M., and Volker, S.
Patterson, G.H., Knobel, S.M., Sharif, W.D., Kain, S.R., and Piston, D.W. 1997.
1999. Three photoconvertible forms of green fluorescent protein identified
Use of the green fluorescent protein and its mutants in quantitative fluores-
by spectral hole-burning. Nat. Struct. Biol. 6: 557–560.
cence microscopy. Biophys. J. 73: 2782–2790.
Creemers, T.M.H., Lock, A.J., Subramaniam, V., Jovin, T.M., and Volker, S.
Phillips, G.N. 1997. Structure and dynamics of green fluorescent protein. Curr.
2000. Photophysics and optical switching in green fluorescent protein mu-
Opin. Struct. Biol. 7: 821–827.
tants. Proc. Natl. Acad. Sci. 97: 2974–2978.
Pierce, D.W., Hom-Booher, N., and Vale R.D. 1997. Imaging individual green
Cubitt, A.B., Heim, R., Adams, S.R., Boyd, A.E., Gross, L.A., and Tsien, R.Y.
fluorescent proteins. Nature 388: 338.
1995. Understanding, improving and using green fluorescent proteins.
Press, W.H., Teukolsky, S.A., Vetterling, W.T., and Flannery, B.P. 1993. Nu-
Trends Biochem. Sci. 20: 448–455.
merical recipes. In The art of scientific computing, 2nd ed. Cambridge
Deschenes, L.A. and Vanden Bout, D.A. 2001. Single-molecule studies of het-
erogeneous dynamics in polymer melts near the class transition. Science
Rigler, R., Mets, U., Widengren, J., and Kask, P. 1993. Fluorescence correlation
292: 255–258.
spectroscopy with high count rate and low background: Analysis of trans-
Diaspro, A., Annunziata, S., Raimondo, M., and Robello, M. 1999a. Three-
lational diffusion. Eur. Biophys. J. 22: 169–175.
dimensional optical behaviour of a confocal microscope with single illumi-
Schwille, P., Kummer, S., Heikal, A.A., Moerner, W.E., and Webb, W.W. 2000.
nation and detection pinhole through imaging of subresolution beads. Mi-
Fluorescence correlation spectroscopy reveals fast optical excitation-driven
crosc. Res. Tech. 45: 130–131
intramolecular dynamics of yellow fluorescent proteins. Proc. Natl. Acad.
Diaspro, A., Corosu, M., Ramoino, P., and Robello, M. 1999b. Adapting a
Sci. 97: 151–156.
compact confocal microscope system to a two-photon excitation fluores-
Shimomura, O., Johnson, F.H., and Saiga, Y. 1962. Extraction, purification and
cence imaging architecture. Microsc. Res. Tech. 47: 196–205.
properties of aequorin, a bioluminescent protein from the luminous hydro-
Dickson, R.M., Cubitt, A., Tsien, R.Y., and Moerner, W.E. 1997. On/off blink-
medusan Aequorea. J. Cell Comp. Physiol. 59: 223–240.
ing and switching behaviour of single molecules of green fluorescent pro-
Song, L., Varma, C.A., Verhoeven, J.W., and Tanke, H.J. 1996. Influence of the
tein. Nature 388: 355–358.
triplet excited state on the photobleaching kinetics of fluorescein in micros-
Edman, L. and Rigler, R. 2000. Memory landscapes of single-enzyme mol-
copy. Biophys. J. 70: 2959–2968.
ecules. Proc. Natl. Acad. Sci. 97: 8266–8271.
Swaminathan, R., Hoang, C.P., and Verkman, A.S. 1997. Photobleaching re-
Ellerby, L.M., Nishida, C.R., Nishida, F., Yamanaka, S.A., Dunn, B., Valentine,
covery and anisotropy decay of green fluorescent protein GFP-S65T in
J.S., Zink, J.I. 1992. Encapsulation of proteins in transparent porous silicate
solution and cells: Cytoplasmic viscosity probed by green fluorescent pro-
glasses prepared by the sol-gel method. Science 255: 1113–1115.
tein translational and rotational diffusion. Biophys. J. 72: 1900–1907.
Elsliger, M.A., Wachter, R.M., Hanson, G.T., Kallio, K., and Remington, S.J.
Talaga, D.S., Lau, W.L., Roder, H., Tang, J., Jia, Y., DeGrado, W., and Hoch-
1999. Structural and spectral response of green fluorescent protein variants
strasser, R.M. 2000. Folding dynamics of single GCN4 peptides by fluo-
to changes in pH. Biochemistry 38: 5296–5301.
rescence resonant energy transfer confocal microscopy. Proc. Natl. Acad.
Ezaz-Nikpay, E., Uchino, K., Lerner, R.E., and Verdine, G.L. 1994. Construc-
Sci. 97: 13021–13026.
tion of an overproduction vector containing the novel srp (sterically re-
Terry, B.R., Matthews, E.K., and Haseloff, J. 1995. Molecular characterization
pressed) promoter. Protein Sci. 3: 132–138.
of recombinant green fluorescent protein by fluorescence correlation mi-
Garcia-Parajo, M.F., Segers-Nolten, G.M.J., Veerman, J.A., Greve, J., and van
croscopy. Biochem. Biophys. Res. Commun. 217: 21–27.
Hulst, N.F. 2000. Real-time light-driven dynamics of the fluorescence emis-
Thompson, R.B. and Gratton, E. 1988. Phase fluorometric method for determi-
sion in single green fluorescent protein molecules. Proc. Natl. Acad. Sci. 97:
nation of standard lifetimes. Anal. Chem. 60: 670–674.
Tsien, R.Y. 1998. The green fluorescent protein. Annu. Rev. Biochem. 67:
Gottfried, D.S., Kagan, A., Hoffman, B.M., and Friedman, J.M. 1999. Impeded
rotation of a protein in a sol-gel matrix. J. Phys. Chem. B. 103: 2803–2807.
Tsien, R. and Wagonner, Y. 1995. Fluorophores for confocal microscopy. In
Haupts, U., Maiti, S., Schwille, P., and Webb, W.W. 1998. Dynamics of fluo-
Handbook of biological confocal microscopy, Vol. 2, (ed. J.B. Pawley), pp.
rescence fluctuations in green fluorescent protein observed by fluorescence
correlation spectroscopy. Proc. Natl. Acad. Sci. 95: 13573–13578.
Volkmer, A., Subramaniam, V., Birch, D.J.S., and Jovin, T.M. 2000. One- and
Heim, R. and Tsien, R.Y. 1996. Engineering green fluorescent protein for im-
two-photon excited fluorescence lifetimes and anisotropy decays of green
proved brightness, longer wavelengths and fluorescence resonance energy
fluorescent proteins. Biophys. J. 78: 1589–1598.
transfer. Curr. Biol. 6: 178–182.
Kelley, A.M., Michalet, X., and Weiss, S. 2001. Single-molecule spectroscopy
Wachter, R.M., Elsliger, M.A., Kallio, K., Hanson, G.T., and Remington, S.J.
comes of age. Science 292: 1671–1672.
1998. Structural basis of spectral shifts in the yellow-emission variants of
Jordan, J.D., Dunbar, R.A., and Bright, F.V. 1995. Dynamics of acrylodan-
green fluorescent protein. Structure 6: 1267–1277.
labeled bovine and human serum albumin entrapped in a sol-gel derived
Weiss, S. 1999. Fluorescence spectroscopy of single biomolecules. Science 283:
biogel. Anal. Chem. 67: 2436–2443.
Lu, H.P., Xun, L., and Xie, X.S. 1998. Single-molecule enzymatic dynamics.
———. 2000. Measuring conformational dynamics of biomolecules by single
Science 282: 1877–1882.
molecule fluorescence spectroscopy. Nat. Struct. Biol. 7: 754–729.
Mozzarelli, A. and Bettati, S. 2001. Functional properties of immobilized pro-
Yang, F., Moss, L.G., and Phillips, G.N. 1996. The molecular structure of green
teins. In Advanced functional molecules and polymers (ed. H.S. Nalwa),
fluorescent protein. Nat. Biotechnol. 14: 1246–1251.
Vol. 4, pp 55–97. Gordon and Breach Science Publishers Association, Sin-
Zhuang, X., Bartley, L.E., Babcock, H.P., Russel, R., Ha, T., Herschlag, D., and
Chu, S. 2000a. A single-molecule study of RNA catalysis and folding.
Ormo, M., Cubitt, A., Kallio, K., Gross, L., Tsien, R.Y., and Remington, S. Science 288: 2048–2051.
1996. Crystal structure of the Aequorea victoria green fluorescent protein.
Zhuang, X., Ha, T., Kim, H.D., Centner, T., Labeit, S., and Chu, S. 2000b. Science 273: 1392–1395.
Fluorescence quenching: A tool for single-molecule protein-folding study.
Palm, G.J., Zdanov, A., Gaitanaris, G.A., Stauber, R., Pavlakis, G.N., and Wlo-
Proc. Natl. Acad. Sci. 97: 14241–14244.
dawer, A. 1997. The structural basis for spectral variations in green fluo-
Zumbusch, A. and Jung, G. 2000. Single molecule spectroscopy of the green
rescent protein. Nat. Struct. Biol. 4: 361–365.
fluorescent protein: A critical assessment. Single Mol. 1: 261–270. www.proteinscience.org
Diário da República, 1.ª série — N.º 166 — 29 de agosto de 2013 ASSEMBLEIA DA REPÚBLICA ções Públicas, aprovado em anexo à Lei n.º 59/2008, de 11 de setembro, alterada pela Lei n.º 3 -B/2010, de 28 de Lei n.º 68/2013 abril, pelo Decreto -Lei n.º 124/2010, de 17 de novembro, e pelas Leis n.os 64 -B/2011, de 30 de dezembro, e 66/2012, de 29 de agosto de 31 de dezem
Hong Kong Exchanges and Clearing Limited and The Stock Exchange of Hong Kong Limitedtake no responsibility for the contents of this announcement, make no representation as to itsaccuracy or completeness and expressly disclaim any liability whatsoever for any losshowsoever arising from or in reliance upon the whole or any part of the contents of thisannouncement. ASIA CEMENT (CHINA) HOLDINGS COR
 Chirico et al.
Chirico et al.
 Spectroscopic properties of immobilized GFPmut2
Spectroscopic properties of immobilized GFPmut2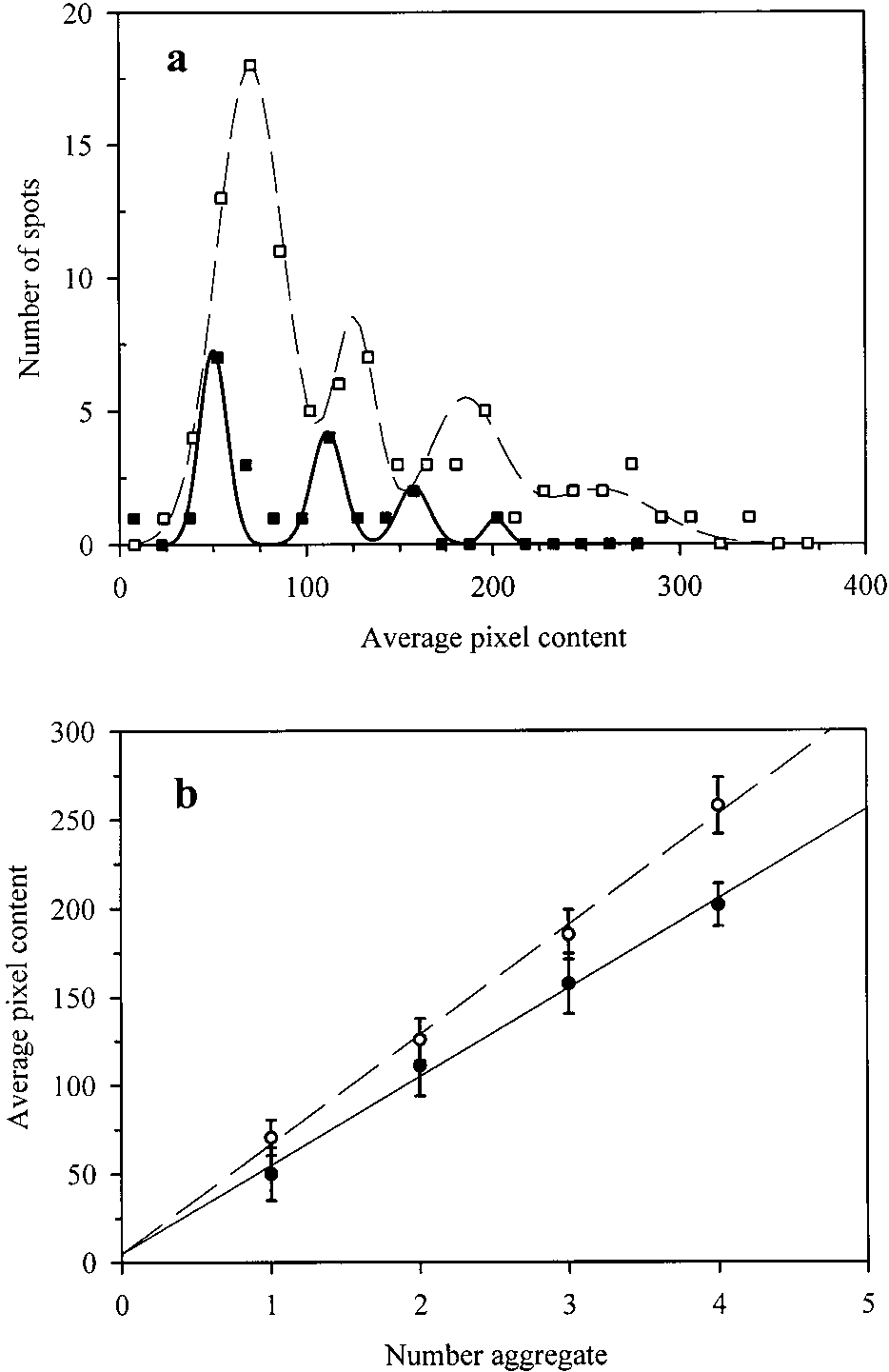
 Chirico et al.
Chirico et al.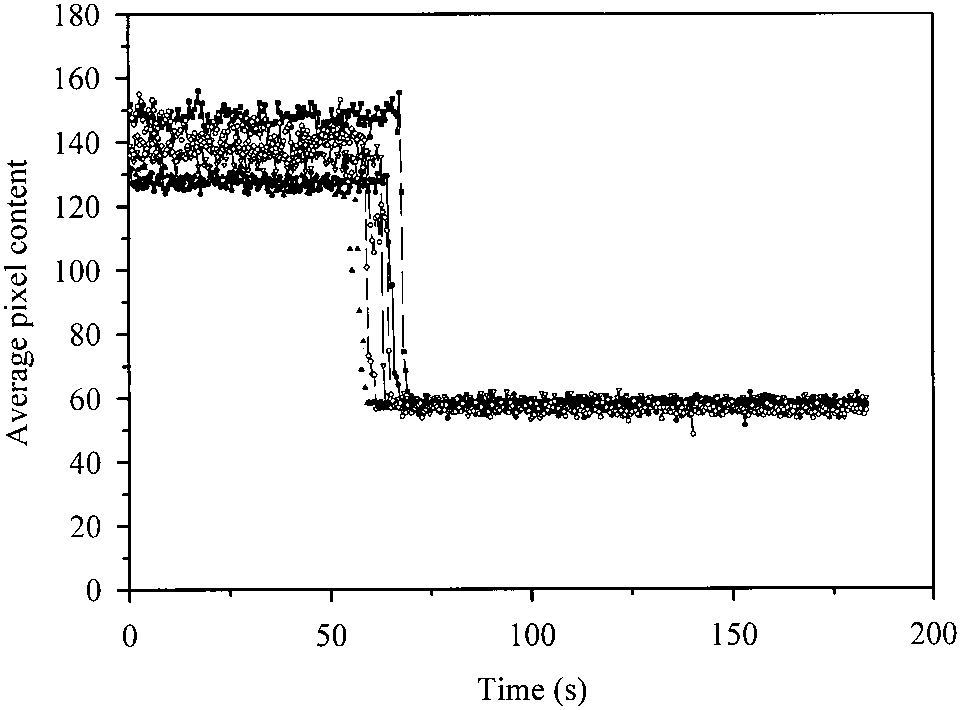
 Spectroscopic properties of immobilized GFPmut2
Spectroscopic properties of immobilized GFPmut2